រចនាសម្ព័ន្ធបន្ទាប់បន្សំនៃ RNA ។ រចនាសម្ព័ន្ធនិងមុខងារនៃអាស៊ីត ribonucleic
ម៉ូលេគុល RNA ក៏ជាវត្ថុធាតុ polymer ដែលម៉ូណូម័រគឺជា ribonucleotides ដែរ RNA គឺជាម៉ូលេគុលខ្សែតែមួយ។ វាត្រូវបានបង្កើតឡើងតាមរបៀបដូចគ្នានឹងខ្សែ DNA មួយ។ នុយក្លេអូទីត RNA គឺស្រដៀងទៅនឹង DNA nucleotides ទោះបីជាមិនដូចគ្នាទៅនឹងពួកវាក៏ដោយ។ វាក៏មាន 4 ក្នុងចំនោមពួកវាដែរ ហើយពួកវាមានសំណល់មូលដ្ឋានអាសូត ផេនតូស និងអាស៊ីតផូស្វ័រ។ មូលដ្ឋានអាសូតទាំងបីគឺដូចគ្នាបេះបិទដូចនៅក្នុង DNA៖ ក, ជីនិង គ. ទោះយ៉ាងណាក៏ដោយផ្ទុយទៅវិញ ធ DNA នៅក្នុង RNA មានមូលដ្ឋាន pyrimidine ដែលមានរចនាសម្ព័ន្ធស្រដៀងគ្នា - uracil ( យូ) ភាពខុសគ្នាសំខាន់រវាង DNA និង RNA គឺជាធម្មជាតិនៃកាបូអ៊ីដ្រាត៖ នៅក្នុង DNA nucleotides monosaccharide គឺ deoxyribose ហើយនៅក្នុង RNA វាគឺជា ribose ។ ការតភ្ជាប់រវាងនុយក្លេអូទីតត្រូវបានអនុវត្តដូចនៅក្នុង DNA តាមរយៈជាតិស្ករ និងសំណល់អាស៊ីតផូស្វ័រ។ មិនដូច DNA ទេ មាតិកាដែលថេរនៅក្នុងកោសិកានៃសារពាង្គកាយមួយចំនួន មាតិកានៃ RNA នៅក្នុងពួកវាប្រែប្រួល។ វាខ្ពស់ជាងគួរឱ្យកត់សម្គាល់ដែលការសំយោគខ្លាំងកើតឡើង។
ទាក់ទងទៅនឹងមុខងារដែលពួកគេអនុវត្ត ប្រភេទ RNA ជាច្រើនត្រូវបានសម្គាល់។
ផ្ទេរ RNA (tRNA) ។ ម៉ូលេគុល tRNA គឺខ្លីបំផុត: ពួកវាមាននុយក្លេអូទីតត្រឹមតែ 80-100 ប៉ុណ្ណោះ។ ទំងន់ម៉ូលេគុលនៃភាគល្អិតបែបនេះគឺ 25-30 ពាន់ RNAs ផ្ទេរត្រូវបានផ្ទុកជាចម្បងនៅក្នុង cytoplasm នៃកោសិកា។ មុខងាររបស់ពួកគេគឺផ្ទេរអាស៊ីតអាមីណូទៅ ribosomes ទៅកាន់កន្លែងសំយោគប្រូតេអ៊ីន។ នៃមាតិកា RNA សរុបនៃកោសិកា tRNA មានប្រហែល 10% ។
រីបូសូម RNA (rRNA) ។ ទាំងនេះគឺជាម៉ូលេគុលធំ៖ ពួកវាមាននុយក្លេអូទីតពី 3 ទៅ 5 ពាន់ រៀងគ្នា ទម្ងន់ម៉ូលេគុលរបស់ពួកគេឡើងដល់ 1-1.5 លាន។ Ribosomal RNAs បង្កើតបានជាផ្នែកសំខាន់នៃ ribosome ។ នៃមាតិកា RNA សរុបនៅក្នុងក្រឡាមួយ rRNA មានប្រហែល 90% ។
កម្មវិធីផ្ញើសារ RNA (mRNA) ឬ អ្នកនាំសារ RNA (mRNA) ត្រូវបានរកឃើញនៅក្នុងស្នូល និង cytoplasm ។ មុខងាររបស់វាគឺដើម្បីផ្ទេរព័ត៌មានអំពីរចនាសម្ព័ន្ធនៃប្រូតេអ៊ីនពី DNA ទៅកាន់ទីតាំងនៃការសំយោគប្រូតេអ៊ីននៅក្នុង ribosomes ។ mRNA មានចំនួនប្រហែល 0.5-1% នៃមាតិកា RNA សរុបនៃកោសិកា។ ទំហំនៃ mRNA ប្រែប្រួលយ៉ាងទូលំទូលាយ - ពី 100 ទៅ 10,000 nucleotides ។
គ្រប់ប្រភេទនៃ RNA ត្រូវបានសំយោគនៅលើ DNA ដែលបម្រើជាទម្រង់គំរូមួយ។
DNA គឺជាអ្នកបញ្ជូនព័ត៌មានតំណពូជ។
ប្រូតេអ៊ីននីមួយៗត្រូវបានតំណាងដោយខ្សែសង្វាក់ polypeptide មួយ ឬច្រើន។ ផ្នែកមួយនៃ DNA ដែលផ្ទុកព័ត៌មានអំពីខ្សែសង្វាក់ polypeptide មួយត្រូវបានគេហៅថា ហ្សែន. ចំនួនសរុបនៃម៉ូលេគុល DNA នៅក្នុងកោសិកាមួយដើរតួជាក្រុមហ៊ុនបញ្ជូនព័ត៌មានហ្សែន។ ព័ត៌មានហ្សែនត្រូវបានបញ្ជូនទាំងពីកោសិកាម្តាយទៅកោសិកាកូនស្រី និងពីឪពុកម្តាយទៅកូន។ ហ្សែនគឺជាអង្គភាពហ្សែន, ឬ ព័ត៌មានតំណពូជ។
DNA គឺជាអ្នកបញ្ជូនព័ត៌មានហ្សែននៅក្នុងកោសិកាមួយ។ - មិនចូលរួមដោយផ្ទាល់ក្នុងការសំយោគប្រូតេអ៊ីន។ នៅក្នុងកោសិកា eukaryotic ម៉ូលេគុល DNA មាននៅក្នុងក្រូម៉ូសូមនៃស្នូល ហើយត្រូវបានបំបែកដោយស្រោមសំបុត្រនុយក្លេអ៊ែរពី cytoplasm ដែលការសំយោគប្រូតេអ៊ីនកើតឡើង។ សារបញ្ជូនព័ត៌មានត្រូវបានបញ្ជូនពីស្នូលទៅកាន់ ribosomes ដែលជាកន្លែងប្រមូលផ្តុំប្រូតេអ៊ីន ហើយអាចឆ្លងកាត់រន្ធញើសនៃភ្នាសនុយក្លេអ៊ែរ។ អ្នកនាំសារនេះគឺជាអ្នកនាំសារ RNA (mRNA) ។ យោងតាមគោលការណ៍នៃការបំពេញបន្ថែម វាត្រូវបានសំយោគនៅលើ DNA ដោយមានការចូលរួមពីអង់ស៊ីមមួយឈ្មោះថា RNA ។ ប៉ូលីមេរ៉ាស.

Messenger RNA គឺជាម៉ូលេគុលដែលមានខ្សែតែមួយ ហើយការចម្លងកើតឡើងពីខ្សែតែមួយនៃម៉ូលេគុល DNA ពីរខ្សែ។ វាមិនមែនជាការចម្លងនៃម៉ូលេគុល DNA ទាំងមូលនោះទេ ប៉ុន្តែមានតែផ្នែកមួយរបស់វាប៉ុណ្ណោះ - ហ្សែនមួយនៅក្នុង eukaryotes ឬក្រុមនៃហ្សែនដែលនៅជាប់គ្នាដែលផ្ទុកព័ត៌មានអំពីរចនាសម្ព័ន្ធនៃប្រូតេអ៊ីនដែលចាំបាច់ដើម្បីអនុវត្តមុខងារមួយនៅក្នុង prokaryotes ។ ក្រុមហ្សែននេះត្រូវបានគេហៅថា operon. នៅដើមនៃ operon នីមួយៗមានប្រភេទនៃបន្ទះចុះចតសម្រាប់ RNA polymerase ហៅថា អ្នកផ្សព្វផ្សាយ.នេះគឺជាលំដាប់ជាក់លាក់នៃ DNA nucleotides ដែលអង់ស៊ីម "ទទួលស្គាល់" ដោយសារតែភាពស្និទ្ធស្នាលគីមី។ មានតែការភ្ជាប់ទៅអ្នកផ្សព្វផ្សាយប៉ុណ្ណោះ RNA polymerase អាចចាប់ផ្តើមសំយោគ RNA ។ ដោយបានឈានដល់ចុងបញ្ចប់នៃ operon អង់ស៊ីមជួបប្រទះនឹងសញ្ញាមួយ (ក្នុងទម្រង់ជាលំដាប់នុយក្លេអូទីតជាក់លាក់) ដែលបង្ហាញពីការបញ្ចប់នៃការអាន។ mRNA ដែលបានបញ្ចប់ចាកចេញពី DNA ហើយទៅកន្លែងសំយោគប្រូតេអ៊ីន។
ដំណើរការចម្លងមានបួនដំណាក់កាល៖ ១) ការភ្ជាប់ RNA-polymerase ជាមួយអ្នកផ្សព្វផ្សាយ; 2) ការចាប់ផ្តើម- ការចាប់ផ្តើមនៃការសំយោគ។ វាមាននៅក្នុងការបង្កើតចំណង phosphodiester ដំបូងរវាង ATP ឬ GTP និង nucleotide ទីពីរនៃម៉ូលេគុល RNA សំយោគ។ 3) ការពន្លូត- ការលូតលាស់នៃខ្សែសង្វាក់ RNA; ទាំងនោះ។ ការបន្ថែមនុយក្លេអូទីតជាបន្តបន្ទាប់ទៅគ្នាទៅវិញទៅមកតាមលំដាប់លំដោយដែលនុយក្លេអូទីតបំពេញបន្ថែមរបស់ពួកគេលេចឡើងនៅក្នុងខ្សែ DNA ដែលបានចម្លង។ អត្រានៃការពន្លូតគឺ 50 nucleotides ក្នុងមួយវិនាទី; ៤) ការបញ្ចប់- បញ្ចប់ការសំយោគ RNA ។
ដោយបានឆ្លងកាត់រន្ធញើសនៃភ្នាសនុយក្លេអ៊ែរ mRNA ត្រូវបានបញ្ជូនទៅ ribosomes ដែលព័ត៌មានហ្សែនត្រូវបានបកស្រាយ - បកប្រែពី "ភាសា" នៃ nucleotides ទៅ "ភាសា" នៃអាស៊ីតអាមីណូ។ ការសំយោគខ្សែសង្វាក់ polypeptide ដោយប្រើម៉ាទ្រីស mRNA ដែលកើតឡើងនៅក្នុង ribosomes ត្រូវបានគេហៅថា ការចាក់ផ្សាយ(ការបកប្រែឡាតាំង - ការបកប្រែ) ។
អាស៊ីតអាមីណូ ដែលប្រូតេអ៊ីនត្រូវបានសំយោគត្រូវបានបញ្ជូនទៅ ribosomes ដោយប្រើ RNAs ពិសេសដែលហៅថា transfer RNAs (tRNAs)។ មាន tRNAs ផ្សេងៗគ្នាជាច្រើននៅក្នុងកោសិកាមួយ ដោយសារមាន codons ដែលសរសេរកូដសម្រាប់អាស៊ីតអាមីណូ។ នៅផ្នែកខាងលើនៃ "ស្លឹក" នៃ tRNA នីមួយៗមានលំដាប់នៃនុយក្លេអូទីតចំនួនបីដែលបំពេញបន្ថែមទៅនឹងនុយក្លេអូទីតនៃ codon នៅក្នុង mRNA ។ ពួកគេហៅនាង ថ្នាំ anticodon ។អង់ស៊ីមពិសេស codase ទទួលស្គាល់ tRNA និងភ្ជាប់អាស៊ីតអាមីណូទៅនឹង "ស្លឹកស្លឹក" - តែមួយគត់ដែលត្រូវបានអ៊ិនកូដដោយ triplet បំពេញបន្ថែមទៅនឹង anticodon ។ ការបង្កើតចំណង covalent រវាង tRNA និងអាស៊ីតអាមីណូ "ផ្ទាល់" របស់វាទាមទារថាមពលនៃម៉ូលេគុល ATP មួយ។
 ដើម្បីឱ្យអាស៊ីតអាមីណូត្រូវបានរួមបញ្ចូលនៅក្នុងខ្សែសង្វាក់ polypeptide វាត្រូវតែបំបែកចេញពី tRNA ។ វាអាចទៅរួចនៅពេលដែល tRNA ចូលទៅក្នុង ribosome ហើយ anticodon ទទួលស្គាល់ codon របស់វានៅក្នុង mRNA ។ ribosome មានទីតាំងពីរសម្រាប់ភ្ជាប់ម៉ូលេគុល tRNA ពីរ។ នៅក្នុងតំបន់មួយក្នុងចំណោមតំបន់ទាំងនេះត្រូវបានគេហៅថា អ្នកទទួល, tRNA មកដល់ជាមួយនឹងអាស៊ីតអាមីណូ ហើយភ្ជាប់ទៅនឹង codon (I) របស់វា។ តើអាស៊ីតអាមីណូនេះភ្ជាប់ទៅនឹងខ្លួនវា (ទទួលយក) ខ្សែសង្វាក់ប្រូតេអ៊ីនដែលកំពុងលូតលាស់ (II) ដែរឬទេ? ចំណង peptide ត្រូវបានបង្កើតឡើងរវាងពួកវា។ tRNA ដែលឥឡូវនេះត្រូវបានភ្ជាប់ជាមួយ mRNA codon នៅក្នុង ម្ចាស់ជំនួយផ្នែកនៃ ribosome ។ tRNA ថ្មីចូលមកក្នុងកន្លែងទទួលទំនេរ ដែលភ្ជាប់ទៅនឹងអាស៊ីតអាមីណូ ដែលត្រូវបានអ៊ិនគ្រីបដោយ codon បន្ទាប់ (III)។ ខ្សែសង្វាក់ polypeptide ដែលបានផ្ដាច់ត្រូវបានផ្ទេរនៅទីនេះម្តងទៀតពីគេហទំព័រម្ចាស់ជំនួយ និងពង្រីកដោយតំណមួយទៀត។ អាស៊ីតអាមីណូនៅក្នុងខ្សែសង្វាក់ដែលកំពុងលូតលាស់ត្រូវបានតភ្ជាប់តាមលំដាប់លំដោយដែល codons អ៊ិនកូដពួកវាមានទីតាំងនៅ mRNA ។
ដើម្បីឱ្យអាស៊ីតអាមីណូត្រូវបានរួមបញ្ចូលនៅក្នុងខ្សែសង្វាក់ polypeptide វាត្រូវតែបំបែកចេញពី tRNA ។ វាអាចទៅរួចនៅពេលដែល tRNA ចូលទៅក្នុង ribosome ហើយ anticodon ទទួលស្គាល់ codon របស់វានៅក្នុង mRNA ។ ribosome មានទីតាំងពីរសម្រាប់ភ្ជាប់ម៉ូលេគុល tRNA ពីរ។ នៅក្នុងតំបន់មួយក្នុងចំណោមតំបន់ទាំងនេះត្រូវបានគេហៅថា អ្នកទទួល, tRNA មកដល់ជាមួយនឹងអាស៊ីតអាមីណូ ហើយភ្ជាប់ទៅនឹង codon (I) របស់វា។ តើអាស៊ីតអាមីណូនេះភ្ជាប់ទៅនឹងខ្លួនវា (ទទួលយក) ខ្សែសង្វាក់ប្រូតេអ៊ីនដែលកំពុងលូតលាស់ (II) ដែរឬទេ? ចំណង peptide ត្រូវបានបង្កើតឡើងរវាងពួកវា។ tRNA ដែលឥឡូវនេះត្រូវបានភ្ជាប់ជាមួយ mRNA codon នៅក្នុង ម្ចាស់ជំនួយផ្នែកនៃ ribosome ។ tRNA ថ្មីចូលមកក្នុងកន្លែងទទួលទំនេរ ដែលភ្ជាប់ទៅនឹងអាស៊ីតអាមីណូ ដែលត្រូវបានអ៊ិនគ្រីបដោយ codon បន្ទាប់ (III)។ ខ្សែសង្វាក់ polypeptide ដែលបានផ្ដាច់ត្រូវបានផ្ទេរនៅទីនេះម្តងទៀតពីគេហទំព័រម្ចាស់ជំនួយ និងពង្រីកដោយតំណមួយទៀត។ អាស៊ីតអាមីណូនៅក្នុងខ្សែសង្វាក់ដែលកំពុងលូតលាស់ត្រូវបានតភ្ជាប់តាមលំដាប់លំដោយដែល codons អ៊ិនកូដពួកវាមានទីតាំងនៅ mRNA ។
នៅពេលមួយក្នុងចំនោមបីបីលេចឡើងនៅលើ ribosome ( UAA, UAG, UGA) ដែលជា "សញ្ញាវណ្ណយុត្តិ" រវាងហ្សែន គ្មាន tRNA ណាមួយអាចយកកន្លែងនៅក្នុងកន្លែងទទួលយកបានទេ។ ការពិតគឺថាមិនមាន anticodons បំពេញបន្ថែមទៅនឹងលំដាប់ nucleotide នៃ "សញ្ញាវណ្ណយុត្តិ" ទេ។ ខ្សែដែលបានផ្ដាច់មិនមានអ្វីភ្ជាប់ទៅកន្លែងទទួលទេ ហើយវាទុក ribosome ។ ការសំយោគប្រូតេអ៊ីនបានបញ្ចប់។
នៅក្នុង prokaryotes ការសំយោគប្រូតេអ៊ីនចាប់ផ្តើមដោយ codon សីហាដែលមានទីតាំងនៅកន្លែងដំបូងក្នុងច្បាប់ចម្លងនៃហ្សែននីមួយៗ កាន់កាប់ទីតាំងបែបនេះនៅក្នុង ribosome ដែល anticodon នៃ tRNA ពិសេសដែលភ្ជាប់ទៅនឹងវាមានអន្តរកម្មជាមួយ សារធាតុ formylmentionine. ទម្រង់ដែលបានកែប្រែនៃអាស៊ីតអាមីណូ methionine ភ្លាមៗចូលទៅក្នុងកន្លែងផ្តល់ជំនួយ ហើយដើរតួជាអក្សរធំនៅក្នុងឃ្លា - ការសំយោគនៃខ្សែសង្វាក់ polypeptide ណាមួយចាប់ផ្តើមជាមួយវានៅក្នុងកោសិកាបាក់តេរី។ ពេលបីដង សីហាមិនមែននៅក្នុងកន្លែងដំបូងទេ ប៉ុន្តែនៅក្នុងច្បាប់ចម្លងនៃហ្សែន វាអ៊ិនកូដអាស៊ីតអាមីណូ methionine ។ បន្ទាប់ពីបញ្ចប់ការសំយោគនៃខ្សែសង្វាក់ polypeptide, formylmethionine ត្រូវបានកាត់ចេញពីវា ហើយអវត្តមានពីប្រូតេអ៊ីនដែលបានបញ្ចប់។
 ដើម្បីបង្កើនការផលិតប្រូតេអ៊ីន mRNA ជារឿយៗឆ្លងកាត់មិនមែនមួយទេប៉ុន្តែ ribosome ជាច្រើនក្នុងពេលតែមួយ។ រចនាសម្ព័ន្ធនេះដែលរួបរួមដោយម៉ូលេគុល mRNA មួយត្រូវបានគេហៅថា ប៉ូលីសូម. ribosome នីមួយៗនៅក្នុងខ្សែក្រវ៉ាត់ conveyor ដូច bead នេះសំយោគប្រូតេអ៊ីនដូចគ្នា។
ដើម្បីបង្កើនការផលិតប្រូតេអ៊ីន mRNA ជារឿយៗឆ្លងកាត់មិនមែនមួយទេប៉ុន្តែ ribosome ជាច្រើនក្នុងពេលតែមួយ។ រចនាសម្ព័ន្ធនេះដែលរួបរួមដោយម៉ូលេគុល mRNA មួយត្រូវបានគេហៅថា ប៉ូលីសូម. ribosome នីមួយៗនៅក្នុងខ្សែក្រវ៉ាត់ conveyor ដូច bead នេះសំយោគប្រូតេអ៊ីនដូចគ្នា។
អាស៊ីតអាមីណូត្រូវបានផ្គត់ផ្គង់ជាបន្តបន្ទាប់ទៅ ribosomes ដោយប្រើ tRNA ។ ដោយបានបរិច្ចាកអាស៊ីតអាមីណូ tRNA ទុក ribosome ហើយភ្ជាប់គ្នាដោយជំនួយពី codase ។ ភាពស៊ីសង្វាក់គ្នាខ្ពស់នៃ "សេវាកម្មរបស់រុក្ខជាតិ" ទាំងអស់សម្រាប់ការផលិតប្រូតេអ៊ីនធ្វើឱ្យវាអាចសំយោគខ្សែសង្វាក់ polypeptide ដែលមានអាស៊ីតអាមីណូរាប់រយក្នុងរយៈពេលពីរបីវិនាទី។
លក្ខណៈសម្បត្តិនៃកូដហ្សែន។ សូមអរគុណដល់ដំណើរការនៃការចម្លងនៅក្នុងកោសិកា ព័ត៌មានត្រូវបានផ្ទេរពី DNA ទៅប្រូតេអ៊ីន
DNA → mRNA → ប្រូតេអ៊ីន
ព័ត៌មានហ្សែនដែលមាននៅក្នុង DNA និង mRNA មាននៅក្នុងលំដាប់នៃនុយក្លេអូទីតនៅក្នុងម៉ូលេគុល។
 តើព័ត៌មានត្រូវបានផ្ទេរពី "ភាសា" នៃនុយក្លេអូទីតទៅជា "ភាសា" នៃអាស៊ីតអាមីណូយ៉ាងដូចម្តេច? ការបកប្រែនេះត្រូវបានអនុវត្តដោយប្រើកូដហ្សែន។ លេខកូដឬលេខកូដគឺជាប្រព័ន្ធនៃនិមិត្តសញ្ញាសម្រាប់បកប្រែទម្រង់នៃព័ត៌មានមួយទៅទម្រង់មួយទៀត។ កូដហ្សែនគឺជាប្រព័ន្ធសម្រាប់កត់ត្រាព័ត៌មានអំពីលំដាប់នៃអាស៊ីតអាមីណូនៅក្នុងប្រូតេអ៊ីនដោយប្រើលំដាប់នៃនុយក្លេអូទីតនៅក្នុង mRNA ។
តើព័ត៌មានត្រូវបានផ្ទេរពី "ភាសា" នៃនុយក្លេអូទីតទៅជា "ភាសា" នៃអាស៊ីតអាមីណូយ៉ាងដូចម្តេច? ការបកប្រែនេះត្រូវបានអនុវត្តដោយប្រើកូដហ្សែន។ លេខកូដឬលេខកូដគឺជាប្រព័ន្ធនៃនិមិត្តសញ្ញាសម្រាប់បកប្រែទម្រង់នៃព័ត៌មានមួយទៅទម្រង់មួយទៀត។ កូដហ្សែនគឺជាប្រព័ន្ធសម្រាប់កត់ត្រាព័ត៌មានអំពីលំដាប់នៃអាស៊ីតអាមីណូនៅក្នុងប្រូតេអ៊ីនដោយប្រើលំដាប់នៃនុយក្លេអូទីតនៅក្នុង mRNA ។
តើកូដហ្សែនមានលក្ខណៈសម្បត្តិអ្វីខ្លះ?
លេខកូដគឺបីដង. RNA មាននុយក្លេអូទីតចំនួនបួន៖ A, G, C, U.ប្រសិនបើយើងព្យាយាមកំណត់អាស៊ីតអាមីណូមួយជាមួយនុយក្លេអូទីតមួយ នោះអាស៊ីតអាមីណូចំនួន 16 ក្នុងចំណោម 20 នឹងនៅតែមិនត្រូវបានសរសេរកូដ។ លេខកូដពីរអក្សរនឹងអ៊ិនគ្រីបអាស៊ីតអាមីណូ 16 ។ ធម្មជាតិបានបង្កើតកូដអក្សរបី ឬបី។ វាមានន័យថា អាស៊ីតអាមីណូ 20 នីមួយៗត្រូវបានអ៊ិនកូដដោយលំដាប់នៃនុយក្លេអូទីតចំនួនបីហៅថា triplet ឬ codon ។
កូដគឺខូច។វាមានន័យថា អាស៊ីតអាមីណូនីមួយៗត្រូវបានសរសេរកូដដោយ codon ច្រើនជាងមួយ។ករណីលើកលែង៖ meteonine និង tryptophan ដែលនីមួយៗត្រូវបានអ៊ិនកូដដោយ triplet មួយ។
លេខកូដគឺច្បាស់។ កូដ codon នីមួយៗសម្រាប់តែអាស៊ីតអាមីណូមួយប៉ុណ្ណោះ។
មាន "សញ្ញាវណ្ណយុត្តិ" រវាងហ្សែន។នៅក្នុងអត្ថបទដែលបានបោះពុម្ព មានរយៈពេលមួយនៅចុងបញ្ចប់នៃឃ្លានីមួយៗ។ ឃ្លាដែលពាក់ព័ន្ធជាច្រើនបង្កើតជាកថាខណ្ឌ។ នៅក្នុងភាសានៃព័ត៌មានហ្សែន កថាខណ្ឌបែបនេះគឺជា operon និង mRNA បំពេញបន្ថែមរបស់វា។ ហ្សែននីមួយៗនៅក្នុង operon prokaryotic ឬហ្សែន eukaryotic ដាច់ដោយឡែកមួយបានអ៊ិនកូដខ្សែសង្វាក់ polypeptide មួយ - ឃ្លាមួយ។ ដោយសារក្នុងករណីខ្លះ ខ្សែសង្វាក់ polypeptide ផ្សេងគ្នាជាច្រើនត្រូវបានបង្កើតជាបន្តបន្ទាប់ពីគំរូ mRNA ពួកគេត្រូវតែបំបែកចេញពីគ្នាទៅវិញទៅមក។ ចំពោះគោលបំណងនេះមានបីដងពិសេសក្នុងឆ្នាំហ្សែន - UAA, UAG, UGA ដែលនីមួយៗបង្ហាញពីការបញ្ចប់នៃការសំយោគនៃសង្វាក់ polypeptide មួយ។ ដូច្នេះ អក្សរបីទាំងនេះមានមុខងារជាសញ្ញាវណ្ណយុត្តិ។ ពួកគេត្រូវបានរកឃើញនៅចុងបញ្ចប់នៃហ្សែននីមួយៗ។
មិនមាន "សញ្ញាវណ្ណយុត្តិ" នៅក្នុងហ្សែនទេ។
កូដគឺមានលក្ខណៈជាសកល។កូដហ្សែនគឺដូចគ្នាសម្រាប់សត្វទាំងអស់ដែលរស់នៅលើផែនដី។ នៅក្នុងបាក់តេរី និងផ្សិត ស្រូវសាលី និងកប្បាស ត្រី និងដង្កូវ កង្កែប និងមនុស្ស លេខបីដូចគ្នាសម្រាប់អាស៊ីតអាមីណូដូចគ្នា។

គោលការណ៍នៃការចម្លង DNA ។ ការបន្តនៃសម្ភារៈហ្សែននៅក្នុងជំនាន់នៃកោសិកា និងសារពាង្គកាយត្រូវបានធានាដោយដំណើរការនេះ។ ការចម្លង - ទ្វេដងនៃម៉ូលេគុល DNA ។ដំណើរការស្មុគ្រស្មាញនេះត្រូវបានអនុវត្តដោយស្មុគ្រស្មាញនៃអង់ស៊ីម និងប្រូតេអ៊ីនជាច្រើនដែលមិនមានសកម្មភាពកាតាលីករ ដែលចាំបាច់ដើម្បីផ្តល់ឱ្យខ្សែសង្វាក់ polynucleotide នូវទម្រង់ដែលចង់បាន។ ជាលទ្ធផលនៃការចម្លង ដុំដែកពីរ DNA ដូចគ្នាបេះបិទត្រូវបានបង្កើតឡើង។ អ្វីដែលគេហៅថាម៉ូលេគុលកូនស្រីទាំងនេះ មិនខុសពីគ្នា ឬពីម៉ូលេគុល DNA របស់ម្តាយដើមឡើយ។ ការចម្លងកើតឡើងនៅក្នុងកោសិកាមុនពេលបែងចែក ដូច្នេះកោសិកាកូនស្រីនីមួយៗទទួលបានម៉ូលេគុល DNA ដូចគ្នាដែលកោសិកាម្តាយមាន។ ដំណើរការចម្លងគឺផ្អែកលើគោលការណ៍មួយចំនួន៖
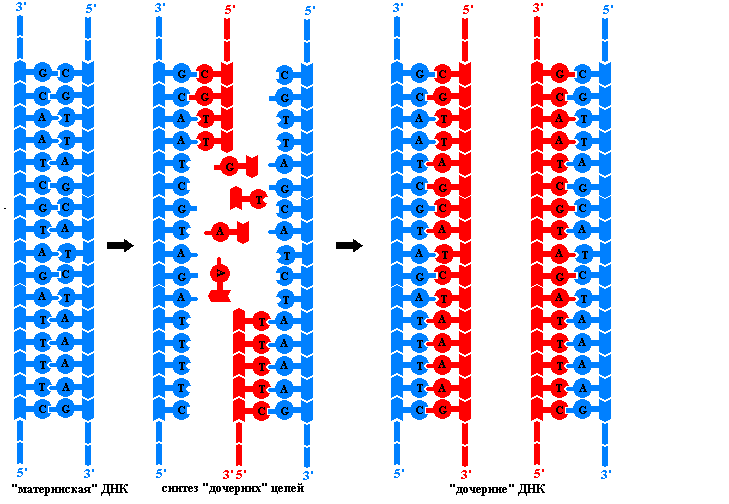
មានតែនៅក្នុងករណីនេះទេដែល DNA polymerases អាចផ្លាស់ទីតាមខ្សែម្តាយ ហើយប្រើវាជាគំរូសម្រាប់ការសំយោគដោយគ្មានកំហុសនៃខ្សែកូនស្រី។ ប៉ុន្តែការពន្លាពេញលេញនៃ helices ដែលមានគូនុយក្លេអូទីតរាប់លានត្រូវបានផ្សារភ្ជាប់ជាមួយនឹងចំនួននៃការបង្វិលដ៏ច្រើនបែបនេះ និងតម្លៃថាមពលដែលមិនអាចទៅរួចទេក្រោមលក្ខខណ្ឌកោសិកា។ ដូច្នេះការចម្លងនៅក្នុង eukaryotes ចាប់ផ្តើមក្នុងពេលដំណាលគ្នានៅកន្លែងខ្លះនៃម៉ូលេគុល DNA ។ តំបន់រវាងចំណុចទាំងពីរដែលការសំយោគនៃច្រវាក់កូនស្រីចាប់ផ្តើមត្រូវបានគេហៅថា ចម្លង. គាត់គឺ ឯកតានៃការចម្លង។

ម៉ូលេគុល DNA នីមួយៗនៃកោសិកា eukaryotic មានផ្ទុកសារចម្លងជាច្រើន។ នៅក្នុងការចម្លងនីមួយៗអ្នកអាចមើលឃើញសមនៃការចម្លង - ផ្នែកនៃម៉ូលេគុល DNA ដែលរលត់រួចហើយនៅក្រោមឥទ្ធិពលនៃអង់ស៊ីមពិសេស។ ខ្សែនីមួយៗនៅក្នុងសមធ្វើជាគំរូសម្រាប់ការសំយោគនៃខ្សែកូនស្រីដែលបំពេញបន្ថែម។ កំឡុងពេលចម្លង ស្តុបផ្លាស់ទីតាមម៉ូលេគុលមេ ហើយផ្នែកថ្មីនៃ DNA បន្ធូរបន្ថយ។ ដោយសារ DNA polymerases អាចផ្លាស់ទីក្នុងទិសដៅតែមួយតាមបណ្តោយខ្សែគំរូ ហើយ strands ត្រូវបានតម្រង់ទិសប្រឆាំងនឹងប៉ារ៉ាឡែល ស្មុគស្មាញអង់ស៊ីមពីរផ្សេងគ្នាត្រូវបានសំយោគក្នុងពេលដំណាលគ្នានៅក្នុងសមនីមួយៗ។ លើសពីនេះទៅទៀត នៅក្នុងសមនីមួយៗ ខ្សែសង្វាក់មួយកូនស្រីម្នាក់ (នាំមុខ) លូតលាស់ជាបន្តបន្ទាប់ ខណៈដែលខ្សែសង្វាក់មួយទៀត (យឺត) ត្រូវបានសំយោគជាបំណែកដាច់ដោយឡែកពីគ្នានូវ nucleotides ជាច្រើនដែលវែង។ អង់ស៊ីមបែបនេះ ត្រូវបានដាក់ឈ្មោះតាមអ្នកវិទ្យាសាស្ត្រជប៉ុន ដែលបានរកឃើញពួកវា បំណែកនៃ Okazakiត្រូវបានភ្ជាប់គ្នាជាមួយ DNA ligase ដើម្បីបង្កើតជាខ្សែសង្វាក់បន្ត។ យន្តការនៃការបង្កើតខ្សែ DNA កូនស្រីដោយបំណែកត្រូវបានគេហៅថាមិនបន្ត។
តម្រូវការសម្រាប់បឋម DNA polymerase មិនអាចផ្តួចផ្តើមការសំយោគនៃខ្សែឈានមុខគេ ឬសំយោគបំណែក Okazaki នៃខ្សែដែលយឺតនោះទេ។ វាអាចពង្រីកខ្សែ polynucleotide ដែលមានស្រាប់ដោយបន្ថែម deoxyribonucleotides ជាបន្តបន្ទាប់ទៅចុង 3'-OH របស់វា។ តើតំបន់ស្ថានីយ 5' ដំបូងនៃខ្សែសង្វាក់ DNA ដែលកំពុងលូតលាស់មកពីណា? វាត្រូវបានសំយោគនៅលើគំរូ DNA ដោយ RNA polymerase ពិសេសហៅថា បឋម(ភាសាអង់គ្លេស Primer - គ្រាប់ពូជ) ។ ទំហំនៃ ribonucleotide primer គឺតូច (តិចជាង 20 nucleotides) បើប្រៀបធៀបទៅនឹងទំហំនៃខ្សែសង្វាក់ DNA ដែលបង្កើតឡើងដោយ DNA poimerase ។ ដោយបានបញ្ចប់នាង មុខងារ ថ្នាំ primer RNA ត្រូវបានយកចេញដោយអង់ស៊ីមពិសេស ហើយគម្លាតដែលបង្កើតឡើងក្នុងអំឡុងពេលដំណើរការនេះត្រូវបានបិទដោយ DNA polymerase ដែលប្រើចុង 3'-OH នៃបំណែក Okazaki ដែលនៅជាប់គ្នាជា primer ។
បញ្ហានៃការចម្លងឡើងវិញនៃចុងនៃម៉ូលេគុល DNA លីនេអ៊ែរ។ ការយកចេញនៃ primers RNA ខ្លាំង, ការបំពេញបន្ថែមទៅនឹងចុង 3' នៃខ្សែទាំងពីរនៃម៉ូលេគុល DNA មេលីនេអ៊ែរ នាំឱ្យការពិតដែលថា strands កូនស្រីខ្លីជាង 10-20 nucleotides ។ នេះគឺជាបញ្ហានៃការចម្លងតិចតួចនៃចុងបញ្ចប់នៃម៉ូលេគុលលីនេអ៊ែរ។
បញ្ហានៃការចម្លងឡើងវិញនៃចុង 3' នៃម៉ូលេគុល DNA លីនេអ៊ែរត្រូវបានដោះស្រាយដោយកោសិកា eukaryotic ដោយប្រើអង់ស៊ីមពិសេស - តេឡូមេរ៉ាស.
Telomerase គឺជា DNA polymerase ដែលបំពេញនូវម៉ូលេគុល DNA 3'-terminal នៃក្រូម៉ូសូមជាមួយនឹងលំដាប់បន្តបន្ទាប់គ្នាខ្លី។ ពួកវាមានទីតាំងនៅខាងក្រោយមួយទៀតបង្កើតរចនាសម្ព័ន្ធស្ថានីយធម្មតារហូតដល់ 10 ពាន់នុយក្លេអូទីតវែង។ បន្ថែមពីលើផ្នែកប្រូតេអ៊ីន telomerase មាន RNA ដែលដើរតួជាគំរូសម្រាប់ពង្រីក DNA ឡើងវិញ។
គ្រោងការណ៍នៃការពន្លូតចុងនៃម៉ូលេគុល DNA ។ ទីមួយ ការភ្ជាប់បន្ថែមនៃចុងដែលលាតសន្ធឹងនៃ DNA ទៅតំបន់គំរូនៃ telomerase RNA កើតឡើង បន្ទាប់មក telomerase ពង្រីក DNA ដោយប្រើចុង 3'-OH របស់វាជា primer ហើយ RNA រួមបញ្ចូលនៅក្នុងអង់ស៊ីមជាគំរូមួយ។ ដំណាក់កាលនេះត្រូវបានគេហៅថាការពន្លូត។ បន្ទាប់ពីនេះ, ការផ្លាស់ប្តូរកើតឡើង, i.e. ចលនានៃ DNA ពង្រីកដោយការធ្វើម្តងទៀតទាក់ទងទៅនឹងអង់ស៊ីម។ នេះត្រូវបានបន្តដោយការពន្លូត និងការប្តូរទីតាំងមួយផ្សេងទៀត។
ជាលទ្ធផលរចនាសម្ព័ន្ធស្ថានីយក្រូម៉ូសូមឯកទេសត្រូវបានបង្កើតឡើង។ ពួកវាមានបណ្តុំ DNA ខ្លីៗម្តងហើយម្តងទៀត និងប្រូតេអ៊ីនជាក់លាក់។
ការសំយោគនៃ rRNA និង tRNA មុនគឺស្រដៀងគ្នាទៅនឹងការសំយោគនៃ ire-mRNA ។ ប្រតិចារិកបឋមនៃ ribosomal RNA មិនមានផ្ទុក introns ហើយនៅក្រោមសកម្មភាពនៃ RNases ជាក់លាក់វាត្រូវបានបំបែកទៅជា 28S-, 18S- និង 5.8S-rRNA; 5S-pRNA ត្រូវបានសំយោគដោយការចូលរួមរបស់ RNA polymerase III ។
rRNA និង tRNA ។
ប្រតិចារឹក tRNA បឋមក៏ត្រូវបានបំប្លែងទៅជាទម្រង់ចាស់ទុំផងដែរដោយផ្នែក hydrolysis ។
គ្រប់ប្រភេទនៃ RNA ត្រូវបានចូលរួមនៅក្នុង biosynthesis ប្រូតេអ៊ីន ប៉ុន្តែមុខងាររបស់ពួកគេនៅក្នុងដំណើរការនេះគឺខុសគ្នា។ តួនាទីរបស់ម៉ាទ្រីសដែលកំណត់រចនាសម្ព័ន្ធចម្បងនៃប្រូតេអ៊ីនត្រូវបានអនុវត្តដោយ messenger RNA (mRNA) ការប្រើប្រាស់ប្រព័ន្ធជីវសំយោគប្រូតេអ៊ីនគ្មានកោសិកាមានសារៈសំខាន់សម្រាប់ការសិក្សាអំពីយន្តការបកប្រែ។ ប្រសិនបើជាលិកា homogenates ត្រូវបាន incubated ជាមួយល្បាយនៃអាស៊ីតអាមីណូ យ៉ាងហោចណាស់មួយដែលត្រូវបានគេដាក់ស្លាក បន្ទាប់មក biosynthesis ប្រូតេអ៊ីនអាចត្រូវបានរកឃើញដោយការបញ្ចូលស្លាកនៅក្នុងប្រូតេអ៊ីន។ រចនាសម្ព័ន្ធចម្បងនៃប្រូតេអ៊ីនដែលត្រូវបានសំយោគត្រូវបានកំណត់ដោយរចនាសម្ព័ន្ធចម្បងនៃ mRNA ដែលបានបន្ថែមទៅក្នុងប្រព័ន្ធ។ ប្រសិនបើប្រព័ន្ធគ្មានកោសិកាត្រូវបានផ្សំឡើងដោយ globin mRNA (វាអាចត្រូវបានញែកចេញពី reticulocytes) globin (a- និង 3-globin chains) ត្រូវបានសំយោគ; ប្រសិនបើ albumin ត្រូវបានសំយោគជាមួយ albumin mRNA ដាច់ដោយឡែកពី hepatocytes ។ល។
១៤. អត្ថន័យចម្លង៖
ក) ដំណើរការគឺជាយន្តការម៉ូលេគុលដ៏សំខាន់ដែលបង្កប់នូវការបែងចែកកោសិកាគ្រប់ប្រភេទនៅក្នុង proeukaryotes ខ) ផ្តល់នូវគ្រប់ប្រភេទនៃការបន្តពូជនៃសារពាង្គកាយឯកតា និងពហុកោសិកា។
គ) រក្សាភាពថេរនៃកោសិកា
សមាសភាពនៃសរីរាង្គ ជាលិកា និងរាងកាយ ជាលទ្ធផលនៃការបង្កើតឡើងវិញខាងសរីរវិទ្យា
ឃ) ធានានូវអត្ថិភាពយូរអង្វែងនៃបុគ្គលជាក់លាក់។
ង) ធានានូវអត្ថិភាពយូរអង្វែងនៃប្រភេទសត្វ។
f) ដំណើរការលើកកម្ពស់ព័ត៌មានត្រឹមត្រូវទ្វេដង។
g) កំហុស (បំរែបំរួល) គឺអាចធ្វើទៅបានក្នុងអំឡុងពេលដំណើរការចម្លងដែលអាចនាំឱ្យមានការរំខានដល់ការសំយោគប្រូតេអ៊ីនជាមួយនឹងការវិវត្តនៃការផ្លាស់ប្តូររោគសាស្ត្រ។
លក្ខណសម្បត្តិពិសេសនៃម៉ូលេគុល DNA ដើម្បីចម្លងខ្លួនវាមុនពេលការបែងចែកកោសិកាត្រូវបានគេហៅថាការចម្លង។
លក្ខណៈសម្បត្តិពិសេសនៃ DNA ដើមជាក្រុមហ៊ុនបញ្ជូនព័ត៌មានតំណពូជ៖
1) ការចម្លង - ការបង្កើតខ្សែសង្វាក់ថ្មីគឺជាការបំពេញបន្ថែម;
2) ការកែតម្រូវដោយខ្លួនឯង - DNA polymerase បំបែកផ្នែកដែលបានចម្លងដោយខុសឆ្គង (10-6);
3) សំណង - ការស្ដារឡើងវិញ;
ដំណើរការទាំងនេះកើតឡើងនៅក្នុងកោសិកាដោយមានការចូលរួមពីអង់ស៊ីមពិសេស។
របៀបដែលប្រព័ន្ធជួសជុលដំណើរការ ការពិសោធន៍ដែលធ្វើឱ្យវាអាចធ្វើទៅបានដើម្បីកំណត់អត្តសញ្ញាណយន្តការនៃការស្ដារឡើងវិញ និងអត្ថិភាពនៃសមត្ថភាពនេះត្រូវបានអនុវត្តដោយប្រើសារពាង្គកាយកោសិកាតែមួយ។ ប៉ុន្តែដំណើរការជួសជុលមាននៅក្នុងកោសិការស់របស់សត្វ និងមនុស្ស។ មនុស្សមួយចំនួនទទួលរងពី xeroderma pigmentosum ។ ជំងឺនេះបណ្តាលមកពីកង្វះសមត្ថភាពកោសិកាក្នុងការសំយោគ DNA ដែលខូចឡើងវិញ។ Xeroderma ត្រូវបានទទួលមរតក។ តើប្រព័ន្ធសំណងមានអ្វីខ្លះ? អង់ស៊ីមចំនួនបួនដែលគាំទ្រដំណើរការជួសជុលគឺ DNA helicase, -exonuclease, -polymerase និង -ligase ។ សមាសធាតុទីមួយនៃសមាសធាតុទាំងនេះអាចទទួលស្គាល់ការខូចខាតនៅក្នុងខ្សែសង្វាក់នៃម៉ូលេគុលអាស៊ីត deoxyribonucleic ។ វាមិនត្រឹមតែទទួលស្គាល់ប៉ុណ្ណោះទេប៉ុន្តែថែមទាំងកាត់ខ្សែសង្វាក់នៅកន្លែងត្រឹមត្រូវដើម្បីដកផ្នែកដែលបានកែប្រែនៃម៉ូលេគុល។ ការលុបបំបាត់ខ្លួនឯងត្រូវបានអនុវត្តដោយប្រើ DNA exonuclease ។ បន្ទាប់មក ផ្នែកថ្មីនៃម៉ូលេគុលអាស៊ីត deoxyribonucleic ត្រូវបានសំយោគពីអាស៊ីតអាមីណូ ដើម្បីជំនួសផ្នែកដែលខូចទាំងស្រុង។ ជាការប្រសើរណាស់, អង្កត់ធ្នូចុងក្រោយនៃនីតិវិធីជីវសាស្រ្តដ៏ស្មុគស្មាញបំផុតនេះត្រូវបានអនុវត្តដោយប្រើអង់ស៊ីម DNA ligase ។ វាទទួលខុសត្រូវចំពោះការភ្ជាប់កន្លែងសំយោគទៅនឹងម៉ូលេគុលដែលខូច។ នៅពេលដែលអង់ស៊ីមទាំងបួនបានធ្វើការងាររបស់ពួកគេ ម៉ូលេគុល DNA ត្រូវបានបន្តឡើងវិញទាំងស្រុង ហើយការខូចខាតទាំងអស់គឺជារឿងអតីតកាល។ នេះជារបៀបដែលយន្តការនៅក្នុងកោសិកាមានជីវិតធ្វើការចុះសម្រុងគ្នា។
ចំណាត់ថ្នាក់នៅពេលនេះ អ្នកវិទ្យាសាស្ត្របែងចែកប្រភេទប្រព័ន្ធសំណងដូចខាងក្រោម។ ពួកវាត្រូវបានធ្វើឱ្យសកម្មអាស្រ័យលើកត្តាផ្សេងៗ។ ទាំងនេះរួមមាន: ការធ្វើឱ្យសកម្មឡើងវិញ។ ការស្តារឡើងវិញ។ ការជួសជុល Heteroduplex ។ ការជួសជុលការកាត់។ ការរួបរួមនៃចុងមិនដូចគ្នានៃម៉ូលេគុល DNA ។ គ្រប់សារពាង្គកាយកោសិកាតែមួយមានប្រព័ន្ធអង់ស៊ីមយ៉ាងតិចបី។ ពួកគេម្នាក់ៗមានសមត្ថភាពអនុវត្តដំណើរការស្តារឡើងវិញ។ ប្រព័ន្ធទាំងនេះរួមមាន: ដោយផ្ទាល់ ការកាត់ចេញ និងក្រោយការចម្លង។ Prokaryotes មានការជួសជុល DNA ទាំងបីប្រភេទនេះ។ ចំពោះ eukaryotes ពួកគេមានយន្តការបន្ថែមដែលហៅថា Miss-mathe និង Sos-repair ។ ជីវវិទ្យាបានសិក្សាយ៉ាងលម្អិតនូវប្រភេទទាំងអស់នៃការព្យាបាលដោយខ្លួនឯងនៃសម្ភារៈហ្សែននៃកោសិកា។
15. កូដហ្សែនគឺជាវិធីសាស្រ្តនៃការអ៊ិនកូដលំដាប់អាស៊ីតអាមីណូនៃប្រូតេអ៊ីនដោយប្រើលំដាប់នៃ nucleotides ដែលជាលក្ខណៈនៃសារពាង្គកាយមានជីវិតទាំងអស់។ លំដាប់អាស៊ីតអាមីណូនៅក្នុងម៉ូលេគុលប្រូតេអ៊ីនត្រូវបានអ៊ិនគ្រីបជាលំដាប់នុយក្លេអូទីតនៅក្នុងម៉ូលេគុល DNA ហើយត្រូវបានគេហៅថា កូដហ្សែន។ផ្នែកនៃម៉ូលេគុល DNA ដែលទទួលខុសត្រូវចំពោះការសំយោគប្រូតេអ៊ីនមួយត្រូវបានគេហៅថា ហ្សែន។
DNA ប្រើនុយក្លេអូទីតចំនួនបួន - adenine (A), guanine (G), cytosine (C), thymine (T) ដែលនៅក្នុងអក្សរសិល្ប៍រុស្ស៊ីត្រូវបានកំណត់ដោយអក្សរ A, G, C និង T ។ អក្សរទាំងនេះបង្កើតជាអក្ខរក្រមនៃ កូដហ្សែន។ RNA ប្រើ nucleotides ដូចគ្នា លើកលែងតែ thymine ដែលត្រូវបានជំនួសដោយ nucleotide ស្រដៀងគ្នា - uracil ដែលត្រូវបានកំណត់ដោយអក្សរ U (U នៅក្នុងអក្សរសិល្ប៍ជាភាសារុស្សី) ។ នៅក្នុងម៉ូលេគុល DNA និង RNA នុយក្លេអូទីតត្រូវបានរៀបចំជាច្រវាក់ ហើយដូច្នេះ លំដាប់នៃអក្សរហ្សែនត្រូវបានទទួល។
ដើម្បីបង្កើតប្រូតេអ៊ីននៅក្នុងធម្មជាតិ អាស៊ីតអាមីណូ 20 ផ្សេងគ្នាត្រូវបានប្រើប្រាស់។ ប្រូតេអ៊ីននីមួយៗគឺជាខ្សែសង្វាក់ ឬខ្សែសង្វាក់ជាច្រើននៃអាស៊ីតអាមីណូនៅក្នុងលំដាប់ដែលបានកំណត់យ៉ាងតឹងរ៉ឹង។ លំដាប់នេះកំណត់រចនាសម្ព័ន្ធនៃប្រូតេអ៊ីន ហើយដូច្នេះលក្ខណៈសម្បត្តិជីវសាស្ត្រទាំងអស់របស់វា។ សំណុំនៃអាស៊ីតអាមីណូក៏ជាសកលសម្រាប់សារពាង្គកាយស្ទើរតែទាំងអស់ផងដែរ។
ការអនុវត្តព័ត៌មានហ្សែននៅក្នុងកោសិការស់ (នោះគឺការសំយោគប្រូតេអ៊ីនដែលបានអ៊ិនកូដដោយហ្សែន) ត្រូវបានអនុវត្តដោយប្រើដំណើរការម៉ាទ្រីសពីរ៖ ការចម្លង (នោះគឺការសំយោគ mRNA នៅលើម៉ាទ្រីស DNA) និងការបកប្រែកូដហ្សែន។ ចូលទៅក្នុងលំដាប់អាស៊ីតអាមីណូ (ការសំយោគនៃខ្សែសង្វាក់ polypeptide នៅលើម៉ាទ្រីស mRNA) ។ នុយក្លេអូទីតបីជាប់ៗគ្នាគឺគ្រប់គ្រាន់ដើម្បីអ៊ិនកូដអាស៊ីតអាមីណូចំនួន 20 ក៏ដូចជាសញ្ញាបញ្ឈប់ដែលបង្ហាញពីការបញ្ចប់នៃលំដាប់ប្រូតេអ៊ីន។ សំណុំនៃនុយក្លេអូទីតបីត្រូវបានគេហៅថា triplet ។ អក្សរកាត់ដែលទទួលយកបានដែលត្រូវគ្នានឹងអាស៊ីតអាមីណូ និងកូដុនត្រូវបានបង្ហាញក្នុងរូប។
លក្ខណៈសម្បត្តិនៃកូដហ្សែន
Triplet - ឯកតានៃកូដដែលមានអត្ថន័យគឺជាការរួមបញ្ចូលគ្នានៃ nucleotides បី (triplet ឬ codon) ។
ការបន្ត - មិនមានសញ្ញាវណ្ណយុត្តិរវាងបីដងទេ ពោលគឺព័ត៌មានត្រូវបានអានជាបន្តបន្ទាប់។
មិនត្រួតស៊ីគ្នា - នុយក្លេអូទីតដូចគ្នាមិនអាចជាផ្នែកនៃកូនបីឬពីរក្នុងពេលដំណាលគ្នា។ (មិនពិតសម្រាប់ហ្សែនត្រួតគ្នាមួយចំនួននៅក្នុងមេរោគ mitochondria និងបាក់តេរីដែលបំប្លែងប្រូតេអ៊ីន frameshift ច្រើន។)
ភាពប្លែក - codon ជាក់លាក់មួយត្រូវគ្នាទៅនឹងអាស៊ីតអាមីណូតែមួយ។ (ទ្រព្យសម្បត្តិមិនមានលក្ខណៈជាសកលទេ។ UGA codon ក្នុង Euplotes crassus អ៊ិនកូដអាស៊ីតអាមីណូពីរ - cysteine និង selenocysteine)
Degeneracy (លែងត្រូវការតទៅទៀត) - codons ជាច្រើនអាចឆ្លើយតបទៅនឹងអាស៊ីតអាមីណូដូចគ្នា។
ភាពជាសកល - កូដហ្សែនដំណើរការដូចគ្នានៅក្នុងសារពាង្គកាយដែលមានកម្រិតផ្សេងៗគ្នានៃភាពស្មុគស្មាញ - ពីមេរោគទៅមនុស្ស (វិធីសាស្រ្តវិស្វកម្មហ្សែនគឺផ្អែកលើនេះ) (មានករណីលើកលែងមួយចំនួនចំពោះទ្រព្យសម្បត្តិនេះ សូមមើលតារាងនៅក្នុង "ការប្រែប្រួលនៃ កូដហ្សែនស្តង់ដារ” ផ្នែកនៅក្នុងអត្ថបទនេះ)។
16.លក្ខខណ្ឌជីវគីមី
ជីវសំយោគប្រូតេអ៊ីនទាមទារព័ត៌មានហ្សែនពីម៉ូលេគុល DNA ។ អ្នកនាំសារ RNA - អ្នកបញ្ជូនព័ត៌មាននេះពីស្នូលទៅកន្លែងសំយោគ; ribosomes - សរីរាង្គដែលការសំយោគប្រូតេអ៊ីនខ្លួនឯងកើតឡើង; សំណុំនៃអាស៊ីតអាមីណូនៅក្នុង cytoplasm; ផ្ទេរ RNAs ដែលអ៊ិនកូដអាស៊ីតអាមីណូនិងផ្ទេរពួកវាទៅកន្លែងនៃការសំយោគនៅលើ ribosomes; ATP គឺជាសារធាតុដែលផ្តល់ថាមពលសម្រាប់ដំណើរការអ៊ិនកូដ និងជីវសំយោគ។
ដំណាក់កាល
ប្រតិចារិក- ដំណើរការនៃជីវសំយោគនៃ RNA គ្រប់ប្រភេទនៅលើម៉ាទ្រីស DNA ដែលកើតឡើងនៅក្នុងស្នូល។
ផ្នែកជាក់លាក់មួយនៃ DNA នៃម៉ូលេគុល despirals ចំណងអ៊ីដ្រូសែនរវាងសង្វាក់ទាំងពីរត្រូវបានបំផ្លាញក្រោមសកម្មភាពរបស់អង់ស៊ីម។ នៅលើខ្សែ DNA មួយ ដូចនៅលើគំរូមួយ ច្បាប់ចម្លង RNA ត្រូវបានសំយោគពី nucleotides តាមគោលការណ៍បំពេញបន្ថែម។ អាស្រ័យលើផ្នែក DNA, ribosomal, transport, and messenger RNAs ត្រូវបានសំយោគតាមរបៀបនេះ។
បន្ទាប់ពីការសំយោគ mRNA វាចាកចេញពីស្នូលហើយត្រូវបានបញ្ជូនទៅ cytoplasm ទៅកន្លែងនៃការសំយោគប្រូតេអ៊ីននៅលើ ribosomes ។
ផ្សាយ- ដំណើរការនៃការសំយោគខ្សែសង្វាក់ polypeptide ធ្វើឡើងនៅលើ ribosomes ដែល mRNA គឺជាអន្តរការីក្នុងការបញ្ជូនព័ត៌មានអំពីរចនាសម្ព័ន្ធចម្បងនៃប្រូតេអ៊ីន។
ជីវសំយោគប្រូតេអ៊ីនមានប្រតិកម្មជាបន្តបន្ទាប់។
1. ការធ្វើឱ្យសកម្ម និងការសរសេរកូដនៃអាស៊ីតអាមីណូ។ tRNA មានរាងដូចស្លឹកឈូក ដែលនៅក្នុងរង្វង់កណ្តាលមានអង់ទីកូដុនបីដង ដែលត្រូវនឹងលេខកូដសម្រាប់អាស៊ីតអាមីណូជាក់លាក់មួយ និង codon នៅលើ mRNA ។ អាស៊ីតអាមីណូនីមួយៗត្រូវបានភ្ជាប់ទៅនឹង tRNA ដែលត្រូវគ្នាដោយប្រើថាមពលរបស់ ATP ។ ស្មុគស្មាញអាស៊ីតអាមីណូ tRNA ត្រូវបានបង្កើតឡើងដែលចូលទៅក្នុង ribosomes ។
2. ការបង្កើតស្មុគស្មាញ mRNA-ribosome ។ mRNA នៅក្នុង cytoplasm ត្រូវបានតភ្ជាប់ដោយ ribosomes នៅលើ granular ER ។
3. ការជួបប្រជុំគ្នានៃខ្សែសង្វាក់ polypeptide ។ tRNA ជាមួយនឹងអាស៊ីតអាមីណូ យោងតាមគោលការណ៍នៃការបំពេញបន្ថែម anticodon-codon រួមបញ្ចូលគ្នាជាមួយ mRNA និងចូលទៅក្នុង ribosome ។ នៅកណ្តាល peptide នៃ ribosome ចំណង peptide ត្រូវបានបង្កើតឡើងរវាងអាស៊ីតអាមីណូពីរ ហើយ tRNA ដែលត្រូវបានបញ្ចេញទុក ribosome ។ ក្នុងករណីនេះ mRNA ឈានទៅមុខមួយដងបីដង ដោយណែនាំ tRNA ថ្មី ដែលជាអាស៊ីតអាមីណូ និងដក tRNA ដែលបញ្ចេញចេញពី ribosome ។ ដំណើរការទាំងមូលត្រូវបានផ្តល់ដោយថាមពល ATP ។ mRNA មួយអាចផ្សំជាមួយ ribosomes ជាច្រើនបង្កើតបានជា polysome ដែលម៉ូលេគុលជាច្រើននៃប្រូតេអ៊ីនមួយត្រូវបានសំយោគក្នុងពេលដំណាលគ្នា។ ការសំយោគបញ្ចប់នៅពេលដែល codons ដែលមិនសមហេតុសមផល (កូដឈប់) ចាប់ផ្តើមនៅលើ mRNA ។ Ribosomes ត្រូវបានបំបែកចេញពី mRNA ហើយខ្សែសង្វាក់ polypeptide ត្រូវបានយកចេញពីពួកគេ។ ចាប់តាំងពីដំណើរការសំយោគទាំងមូលកើតឡើងនៅលើ granular endoplasmic reticulum ច្រវាក់ polypeptide លទ្ធផលចូលទៅក្នុង tubules ER ដែលពួកគេទទួលបានរចនាសម្ព័ន្ធចុងក្រោយរបស់ពួកគេហើយត្រូវបានបំលែងទៅជាម៉ូលេគុលប្រូតេអ៊ីន។
ប្រតិកម្មសំយោគទាំងអស់ត្រូវបានជំរុញដោយអង់ស៊ីមពិសេសជាមួយនឹងការចំណាយថាមពល ATP ។ អត្រានៃការសំយោគគឺខ្ពស់ណាស់ ហើយអាស្រ័យលើប្រវែងនៃសារធាតុ polypeptide ។ ឧទាហរណ៍នៅក្នុង ribosome នៃ Escherichia coli ប្រូតេអ៊ីននៃអាស៊ីតអាមីណូ 300 ត្រូវបានសំយោគក្នុងរយៈពេលប្រហែល 15-20 វិនាទី។
អន្តរកម្មនិងរចនាសម្ព័ន្ធនៃ mRNA, tRNA, RRNA - អាស៊ីត nucleic សំខាន់បី - ត្រូវបានពិចារណាដោយវិទ្យាសាស្រ្តនៃ cytology ។ វានឹងជួយបញ្ជាក់ពីតួនាទីនៃការដឹកជញ្ជូនអាស៊ីត ribonucleic (tRNA) នៅក្នុងកោសិកា។ នេះតូចណាស់ ប៉ុន្តែក្នុងពេលជាមួយគ្នានេះ ម៉ូលេគុលសំខាន់មិនអាចប្រកែកបានចូលរួមក្នុងដំណើរការនៃការផ្សំប្រូតេអ៊ីនដែលបង្កើតជារាងកាយ។
តើរចនាសម្ព័ន្ធរបស់ tRNA គឺជាអ្វី? វាគួរឱ្យចាប់អារម្មណ៍ខ្លាំងណាស់ក្នុងការមើលសារធាតុនេះ "ពីខាងក្នុង" ដើម្បីរកមើលជីវគីមីនិងតួនាទីជីវសាស្រ្តរបស់វា។ ហើយផងដែរ តើរចនាសម្ព័ន្ធរបស់ tRNA និងតួនាទីរបស់វាក្នុងការសំយោគប្រូតេអ៊ីនមានទំនាក់ទំនងគ្នាយ៉ាងដូចម្តេច?
តើ tRNA ជាអ្វី តើវាមានរចនាសម្ព័ន្ធដូចម្តេច?
ការដឹកជញ្ជូនអាស៊ីត ribonucleic ត្រូវបានចូលរួមនៅក្នុងការសាងសង់ប្រូតេអ៊ីនថ្មី។ ស្ទើរតែ 10% នៃអាស៊ីត ribonucleic ទាំងអស់គឺជាមធ្យោបាយដឹកជញ្ជូន។ ដើម្បីបញ្ជាក់ឱ្យច្បាស់ថាតើធាតុគីមីអ្វីខ្លះដែលម៉ូលេគុលត្រូវបានបង្កើតឡើង យើងនឹងរៀបរាប់អំពីរចនាសម្ព័ន្ធនៃរចនាសម្ព័ន្ធបន្ទាប់បន្សំនៃ tRNA ។ រចនាសម្ព័ន្ធបន្ទាប់បន្សំពិចារណាពីចំណងគីមីជាមូលដ្ឋានទាំងអស់រវាងធាតុ។
វាគឺជាម៉ាក្រូម៉ូលេគុលដែលមានខ្សែសង្វាក់ polynucleotide ។ មូលដ្ឋានអាសូតនៅក្នុងវាត្រូវបានតភ្ជាប់ដោយចំណងអ៊ីដ្រូសែន។ ដូច DNA ដែរ RNA មានមូលដ្ឋានអាសូតចំនួន 4៖ អាឌីនីន ស៊ីតូស៊ីន ហ្គានីន និងអ៊ុយរ៉ាស៊ីល។ នៅក្នុងសមាសធាតុទាំងនេះ adenine តែងតែត្រូវបានផ្សារភ្ជាប់ជាមួយនឹង uracil និង guanine ដូចធម្មតាជាមួយ cytosine ។
ហេតុអ្វីបានជានុយក្លេអូទីតមានបុព្វបទ ribo-? សាមញ្ញ ប៉ូលីលីនេអ៊ែរទាំងអស់ដែលមាន ribose ជំនួសឱ្យ pentose នៅមូលដ្ឋាននៃ nucleotide ត្រូវបានគេហៅថា ribonucleic ។ ហើយការផ្ទេរ RNA គឺជាប្រភេទមួយក្នុងចំណោម 3 ប្រភេទនៃវត្ថុធាតុ polymer ribonucleic បែបនេះ។
រចនាសម្ព័ន្ធនៃ tRNA: ជីវគីមី
សូមក្រឡេកមើលស្រទាប់ជ្រៅបំផុតនៃរចនាសម្ព័ន្ធរបស់ម៉ូលេគុល។ នុយក្លេអូទីតទាំងនេះមាន ៣ សមាសធាតុ៖
- Sucrose គ្រប់ប្រភេទនៃ RNA ពាក់ព័ន្ធនឹង ribose ។
- អាស៊ីត phosphoric។
- មូលដ្ឋានអាសូត។ ទាំងនេះគឺជា purines និង pyrimidines ។

មូលដ្ឋានអាសូតត្រូវបានភ្ជាប់ទៅគ្នាទៅវិញទៅមកដោយចំណងដ៏រឹងមាំ។ វាជាទម្លាប់ក្នុងការបែងចែកមូលដ្ឋានទៅជា purine និង pyrimidine ។
Purines គឺ adenine និង guanine ។ Adenine ត្រូវគ្នាទៅនឹង adenyl nucleotide នៃ 2 rings ទាក់ទងគ្នាទៅវិញទៅមក។ ហើយ guanine ត្រូវគ្នាទៅនឹង "ringle-ring" nucleotide guanine ។
Pyramidins គឺជា cytosine និង uracil ។ Pyrimidines មានរចនាសម្ព័ន្ធចិញ្ចៀនតែមួយ។ មិនមាន thymine នៅក្នុង RNA ទេព្រោះវាត្រូវបានជំនួសដោយធាតុដូចជា uracil ។ នេះជាការសំខាន់ក្នុងការយល់ដឹងមុននឹងយកចិត្តទុកដាក់លើលក្ខណៈរចនាសម្ព័ន្ធផ្សេងទៀតនៃ tRNA ។
ប្រភេទនៃ RNA
ដូចដែលអ្នកអាចឃើញរចនាសម្ព័ន្ធនៃ tRNA មិនអាចពិពណ៌នាដោយសង្ខេបបានទេ។ មនុស្សម្នាក់ត្រូវតែស្វែងយល់ពីជីវគីមី ដើម្បីយល់ពីគោលបំណងនៃម៉ូលេគុល និងរចនាសម្ព័ន្ធពិតរបស់វា។ តើ nucleotides ribosomal អ្វីផ្សេងទៀតត្រូវបានគេស្គាល់? ក៏មានម៉ាទ្រីស ឬព័ត៌មាន និងអាស៊ីតនុយក្លេអ៊ីក ribosomal ផងដែរ។ អក្សរកាត់ជា mRNA និង RRNA ។ ម៉ូលេគុលទាំង 3 ធ្វើការយ៉ាងជិតស្និទ្ធជាមួយគ្នានៅក្នុងកោសិកា ដើម្បីធានាថារាងកាយទទួលបានប្រូតេអ៊ីន globules ដែលមានរចនាសម្ព័ន្ធត្រឹមត្រូវ។

វាមិនអាចទៅរួចទេក្នុងការស្រមៃមើលការងាររបស់វត្ថុធាតុ polymer មួយដោយគ្មានជំនួយពី 2 ផ្សេងទៀត។ លក្ខណៈរចនាសម្ព័ន្ធនៃ tRNA កាន់តែច្បាស់នៅពេលពិចារណាដោយភ្ជាប់ជាមួយមុខងារដែលទាក់ទងដោយផ្ទាល់ទៅនឹងការងាររបស់ ribosomes ។
រចនាសម្ព័ន្ធនៃ mRNA, tRNA, និង RRNA គឺស្រដៀងគ្នាតាមវិធីជាច្រើន។ ទាំងអស់មាន ribose នៅមូលដ្ឋានរបស់ពួកគេ។ ទោះជាយ៉ាងណាក៏ដោយរចនាសម្ព័ន្ធនិងមុខងាររបស់ពួកគេគឺខុសគ្នា។
ការរកឃើញអាស៊ីត nucleic
ជនជាតិស្វីស Johann Miescher បានរកឃើញម៉ាក្រូម៉ូលេគុលនៅក្នុងស្នូលកោសិកានៅឆ្នាំ 1868 ដែលក្រោយមកហៅថា នុយក្លេអ៊ែ។ ឈ្មោះ "នុយក្លេអ៊ែរ" មកពីពាក្យ (ស្នូល) - ស្នូល។ ទោះបីជាបន្តិចក្រោយមកគេបានរកឃើញថានៅក្នុងសត្វកោសិកាតែមួយដែលមិនមានស្នូល សារធាតុទាំងនេះក៏មានវត្តមានដែរ។ នៅពាក់កណ្តាលសតវត្សទី 20 គាត់បានទទួលរង្វាន់ណូបែលសម្រាប់ការរកឃើញនៃការសំយោគអាស៊ីត nucleic ។
មុខងាររបស់ tRNA ក្នុងការសំយោគប្រូតេអ៊ីន
ឈ្មោះខ្លួនវា - ផ្ទេរ RNA - និយាយអំពីមុខងារសំខាន់នៃម៉ូលេគុល។ អាស៊ីត nucleic នេះ "នាំ" មកជាមួយវានូវអាស៊ីតអាមីណូសំខាន់ដែលត្រូវការដោយ ribosomal RNA ដើម្បីបង្កើតប្រូតេអ៊ីនជាក់លាក់មួយ។
ម៉ូលេគុល tRNA មានមុខងារតិចតួច។ ទីមួយគឺការទទួលស្គាល់ mRNA codon មុខងារទីពីរគឺការផ្តល់ប្លុកអាគារ - អាស៊ីតអាមីណូសម្រាប់ការសំយោគប្រូតេអ៊ីន។ អ្នកជំនាញខ្លះទៀតគូសបញ្ជាក់ពីមុខងារអ្នកទទួល។ នោះគឺការបន្ថែមអាស៊ីតអាមីណូតាមគោលការណ៍ covalent ។ អង់ស៊ីមដូចជា aminocil-TRNA synthatase ជួយ "ភ្ជាប់" អាស៊ីតអាមីណូនេះ។
តើរចនាសម្ព័ន្ធរបស់ tRNA ទាក់ទងនឹងមុខងាររបស់វាយ៉ាងដូចម្តេច? អាស៊ីត ribonucleic ពិសេសនេះត្រូវបានរចនាឡើងតាមរបៀបដែលនៅម្ខាងមានមូលដ្ឋានអាសូតដែលតែងតែភ្ជាប់ជាគូ។ ទាំងនេះគឺជាធាតុដែលយើងស្គាល់ - A, U, C, G. ពិតប្រាកដ 3 អក្សរឬមូលដ្ឋានអាសូតបង្កើតជា anticodon - សំណុំបញ្ច្រាសនៃធាតុដែលមានអន្តរកម្មជាមួយ codon យោងទៅតាមគោលការណ៍នៃការបំពេញបន្ថែម។
លក្ខណៈសំខាន់នៃរចនាសម្ព័ន្ធ tRNA នេះធានាថានឹងមិនមានកំហុសនៅពេលឌិកូដអាស៊ីត nucleic គំរូ។ យ៉ាងណាមិញ លំដាប់ពិតប្រាកដនៃអាស៊ីតអាមីណូកំណត់ថាតើប្រូតេអ៊ីនដែលរាងកាយត្រូវការនាពេលបច្ចុប្បន្នត្រូវបានសំយោគយ៉ាងត្រឹមត្រូវឬអត់។
លក្ខណៈរចនាសម្ព័ន្ធ
តើលក្ខណៈរចនាសម្ព័ន្ធរបស់ tRNA និងតួនាទីជីវសាស្រ្តរបស់វាមានអ្វីខ្លះ? នេះគឺជារចនាសម្ព័ន្ធបុរាណណាស់។ វិមាត្ររបស់វាស្ថិតនៅចន្លោះ 73 និង 93 nucleotides ។ ទំងន់ម៉ូលេគុលនៃសារធាតុគឺ 25,000-30,000 ។
រចនាសម្ព័ន្ធនៃរចនាសម្ព័ន្ធបន្ទាប់បន្សំនៃ tRNA អាចត្រូវបានវិភាគដោយសិក្សាធាតុសំខាន់ៗចំនួន 5 នៃម៉ូលេគុល។ ដូច្នេះអាស៊ីត nucleic នេះមានធាតុដូចខាងក្រោមៈ
- រង្វិលជុំសម្រាប់ទំនាក់ទំនងជាមួយអង់ស៊ីម;
- រង្វិលជុំសម្រាប់ទំនាក់ទំនងជាមួយ ribosome;
- រង្វិលជុំ anticodon;
- ដើមអ្នកទទួល;
- anticodon ខ្លួនវាផ្ទាល់។
ហើយពួកគេក៏ញែករង្វិលជុំអថេរតូចមួយនៅក្នុងរចនាសម្ព័ន្ធបន្ទាប់បន្សំផងដែរ។ ដៃម្ខាងនៃ tRNA គ្រប់ប្រភេទគឺដូចគ្នា - ដើមនៃសំណល់ cytosine ពីរ និងសំណល់ adenosine មួយ។ វាគឺនៅចំណុចនេះដែលការតភ្ជាប់កើតឡើងជាមួយ 1 នៃអាស៊ីតអាមីណូដែលមាន 20 ។ អាស៊ីតអាមីណូនីមួយៗមានអង់ស៊ីមរបស់វា ដែលជា aminoacyl-tRNA ផ្ទាល់ខ្លួនរបស់វា។

ព័ត៌មានទាំងអស់ដែលអ៊ិនគ្រីបរចនាសម្ព័ន្ធនៃអាស៊ីត nucleic ទាំងអស់មាននៅក្នុង DNA ខ្លួនវាផ្ទាល់។ រចនាសម្ព័ន្ធរបស់ TRNA គឺស្ទើរតែដូចគ្នាបេះបិទនៅក្នុងសត្វមានជីវិតទាំងអស់នៅលើភពផែនដី។ វានឹងមើលទៅដូចស្លឹកឈើពេលមើលជា 2-D។
ទោះជាយ៉ាងណាក៏ដោយ ប្រសិនបើអ្នកមើលបីវិមាត្រ ម៉ូលេគុលនេះប្រហាក់ប្រហែលនឹងរចនាសម្ព័ន្ធធរណីមាត្ររាង L ។ នេះត្រូវបានចាត់ទុកថាជារចនាសម្ព័ន្ធទីបីនៃ tRNA ។ ប៉ុន្តែដើម្បីភាពងាយស្រួលនៃការសិក្សា វាជាទម្លាប់ក្នុងការ "បន្ធូរអារម្មណ៍" វាដោយមើលឃើញ។ រចនាសម្ព័ន្ធទីបីត្រូវបានបង្កើតឡើងជាលទ្ធផលនៃអន្តរកម្មនៃធាតុនៃរចនាសម្ព័ន្ធបន្ទាប់បន្សំដែលជាផ្នែកទាំងនោះដែលបំពេញគ្នាទៅវិញទៅមក។
អាវុធ ឬចិញ្ចៀន TRNA ដើរតួនាទីយ៉ាងសំខាន់។ ជាឧទាហរណ៍ ដៃមួយត្រូវបានទាមទារសម្រាប់ការភ្ជាប់គីមីជាមួយនឹងអង់ស៊ីមជាក់លាក់មួយ។
លក្ខណៈពិសេសមួយនៃ nucleotide គឺវត្តមាននៃ nucleosides មួយចំនួនធំ។ មាន nucleosides តូចៗជាង 60 ប្រភេទ។
រចនាសម្ព័ន្ធ TRNA និងការសរសេរកូដអាស៊ីតអាមីណូ
យើងដឹងថា TRNA anticodon មានប្រវែង 3 ម៉ូលេគុល។ Anticodon នីមួយៗត្រូវគ្នាទៅនឹងអាស៊ីតអាមីណូ "ផ្ទាល់ខ្លួន" ជាក់លាក់។ អាស៊ីតអាមីណូនេះត្រូវបានភ្ជាប់ទៅនឹងម៉ូលេគុល tRNA ដោយប្រើអង់ស៊ីមពិសេស។ ដរាបណាអាស៊ីតអាមីណូ 2 បញ្ចូលគ្នា ចំណងជាមួយ tRNA ត្រូវបានខូច។ សមាសធាតុគីមីនិងអង់ស៊ីមទាំងអស់គឺចាំបាច់មុនពេលវេលាដែលត្រូវការ។ នេះជារបៀបដែលរចនាសម្ព័ន្ធ និងមុខងាររបស់ tRNA មានទំនាក់ទំនងគ្នាទៅវិញទៅមក។
សរុបមក មានម៉ូលេគុលបែបនេះចំនួន 61 ប្រភេទដែលមាននៅក្នុងកោសិកា។ វាអាចមានបំរែបំរួលគណិតវិទ្យាចំនួន 64 ។ ទោះជាយ៉ាងណាក៏ដោយ tRNA 3 ប្រភេទត្រូវបានបាត់ដោយសារតែការពិតដែលថាចំនួនដូចគ្នានៃ stop codons នៅក្នុង mRNA មិនមាន anticodons ។
អន្តរកម្មនៃ mRNA និង tRNA
ចូរយើងពិចារណាពីអន្តរកម្មនៃសារធាតុជាមួយ mRNA និង RRNA ក៏ដូចជាលក្ខណៈរចនាសម្ព័ន្ធនៃ tRNA ។ រចនាសម្ព័ន្ធ និងគោលបំណងនៃម៉ាក្រូម៉ូលេគុលគឺទាក់ទងគ្នាទៅវិញទៅមក។
រចនាសម្ព័ន្ធនៃ mRNA ចម្លងព័ត៌មានពីផ្នែកដាច់ដោយឡែកនៃ DNA ។ DNA ខ្លួនវាធំពេកជាសមាសធាតុនៃម៉ូលេគុល ហើយវាមិនដែលចាកចេញពីស្នូលទេ។ ដូច្នេះយើងត្រូវការ RNA អន្តរការី - ព័ត៌មាន RNA ។

ដោយផ្អែកលើលំដាប់នៃម៉ូលេគុលដែល mRNA បានចម្លង ribosome បង្កើតប្រូតេអ៊ីន។ ribosome គឺជារចនាសម្ព័ន្ធ polynucleotide ដាច់ដោយឡែក ដែលរចនាសម្ព័ន្ធដែលចាំបាច់ត្រូវបញ្ជាក់។
Ribosomal tRNA: អន្តរកម្ម
Ribosomal RNA គឺជាសរីរាង្គដ៏ធំមួយ។ ទម្ងន់ម៉ូលេគុលរបស់វាគឺ 1,000,000 - 1,500,000 ។ស្ទើរតែ 80% នៃបរិមាណ RNA សរុបគឺជា ribosomal nucleotides ។

វាហាក់ដូចជាចាប់យកខ្សែសង្វាក់ mRNA ហើយរង់ចាំ anticodons ដែលនឹងនាំម៉ូលេគុល tRNA ជាមួយពួកគេ។ Ribosomal RNA មាន 2 អនុរង៖ តូច និងធំ។
ribosome ត្រូវបានគេហៅថា "រោងចក្រ" ពីព្រោះការសំយោគសារធាតុទាំងអស់ដែលចាំបាច់សម្រាប់ជីវិតប្រចាំថ្ងៃកើតឡើងនៅក្នុងសរីរាង្គនេះ។ នេះក៏ជារចនាសម្ព័ន្ធកោសិកាបុរាណផងដែរ។
តើការសំយោគប្រូតេអ៊ីនកើតឡើងយ៉ាងដូចម្តេចនៅក្នុង ribosome?
រចនាសម្ព័ន្ធនៃ tRNA និងតួនាទីរបស់វាក្នុងការសំយោគប្រូតេអ៊ីនមានទំនាក់ទំនងគ្នាទៅវិញទៅមក។ anticodon ដែលមានទីតាំងនៅម្ខាងនៃអាស៊ីត ribonucleic មានលក្ខណៈសមរម្យសម្រាប់មុខងារចម្បងរបស់វា - ការផ្តល់អាស៊ីតអាមីណូទៅ ribosome ដែលជាកន្លែងបង្កើតប្រូតេអ៊ីនបន្តិចម្តង ៗ ។ សំខាន់ tRNA ដើរតួជាអន្តរការី។ ភារកិច្ចរបស់វាគឺគ្រាន់តែនាំយកអាស៊ីតអាមីណូចាំបាច់ប៉ុណ្ណោះ។
នៅពេលដែលព័ត៌មានត្រូវបានអានពីផ្នែកមួយនៃ mRNA នោះ ribosome ផ្លាស់ទីបន្ថែមទៀតតាមខ្សែសង្វាក់។ ម៉ាទ្រីសគឺត្រូវការតែដើម្បីបញ្ជូនព័ត៌មានដែលបានអ៊ិនកូដអំពីការកំណត់រចនាសម្ព័ន្ធ និងមុខងារនៃប្រូតេអ៊ីនបុគ្គលប៉ុណ្ណោះ។ បន្ទាប់មក tRNA មួយទៀតដែលមានមូលដ្ឋានអាសូតរបស់វាចូលទៅជិត ribosome ។ វាក៏ឌិកូដផ្នែកបន្ទាប់នៃ mRNA ផងដែរ។
ការឌិកូដកើតឡើងដូចខាងក្រោម។ មូលដ្ឋានអាសូតត្រូវបានរួមបញ្ចូលគ្នាតាមគោលការណ៍នៃការបំពេញបន្ថែមក្នុងវិធីដូចគ្នានឹង DNA ខ្លួនវាដែរ។ ដូច្នោះហើយ TRNA មើលឃើញកន្លែងដែលវាត្រូវការ "moor" និង "hangar" ដើម្បីបញ្ជូនអាស៊ីតអាមីណូ។

បន្ទាប់មកនៅក្នុង ribosome អាស៊ីដអាមីណូដែលត្រូវបានជ្រើសរើសតាមរបៀបនេះត្រូវបានផ្សារភ្ជាប់គ្នាដោយគីមី មួយជំហានម្តងមួយៗ macromolecule លីនេអ៊ែរថ្មីត្រូវបានបង្កើតឡើង ដែលបន្ទាប់ពីការបញ្ចប់ការសំយោគត្រូវបានបង្វិលទៅជា globule (បាល់) ។ tRNA និង mRNA ដែលបានប្រើដោយបានបំពេញមុខងាររបស់ពួកគេត្រូវបានដកចេញពីប្រូតេអ៊ីន "រោងចក្រ" ។
នៅពេលដែលផ្នែកដំបូងនៃ codon គូជាមួយ anticodon នោះស៊ុមអានត្រូវបានកំណត់។ បនា្ទាប់មក ប្រសិនបើដោយសារហេតុផលខ្លះការប្តូរស៊ុមកើតឡើង នោះមុខងារប្រូតេអ៊ីនមួយចំនួននឹងត្រូវបានច្រានចោល។ ribosome មិនអាចធ្វើអន្តរាគមន៍ក្នុងដំណើរការនេះ និងដោះស្រាយបញ្ហាបានទេ។ លុះត្រាតែដំណើរការចប់សព្វគ្រប់ អនុផ្នែក RRNA 2 បញ្ចូលគ្នាម្តងទៀត។ ជាមធ្យម មាន 1 កំហុសសម្រាប់រាល់ 10 4 អាមីណូអាស៊ីត។ សម្រាប់រាល់ប្រូតេអ៊ីន 25 ដែលបានប្រមូលផ្តុំរួចហើយ វាមានកំហុសក្នុងការចម្លងយ៉ាងហោចណាស់ 1 ។
TRNAs ជាម៉ូលេគុលវត្ថុបុរាណ
ចាប់តាំងពី tRNA ប្រហែលជាមាននៅដើមកំណើតនៃជីវិតនៅលើផែនដី វាត្រូវបានគេហៅថាម៉ូលេគុល relict ។ វាត្រូវបានគេជឿថា RNA គឺជារចនាសម្ព័ន្ធដំបូងដែលមានមុន DNA ហើយបន្ទាប់មកមានការវិវត្ត។ សម្មតិកម្មពិភពលោក RNA ត្រូវបានបង្កើតឡើងក្នុងឆ្នាំ 1986 ដោយម្ចាស់ជ័យលាភី Walter Gilbert ។ ទោះជាយ៉ាងណាក៏ដោយនេះនៅតែពិបាកក្នុងការបញ្ជាក់។ ទ្រឹស្ដីនេះត្រូវបានគាំទ្រដោយការពិតជាក់ស្តែង - ម៉ូលេគុល tRNA អាចរក្សាទុកបណ្តុំព័ត៌មាន និងអនុវត្តព័ត៌មាននេះដោយរបៀបណា ពោលគឺអនុវត្តការងារ។
ប៉ុន្តែអ្នកប្រឆាំងទ្រឹស្តីអះអាងថា រយៈពេលខ្លីនៃជីវិតរបស់សារធាតុមិនអាចធានាថា tRNA គឺជាអ្នកបញ្ជូនព័ត៌មានជីវសាស្រ្តល្អណាមួយឡើយ។ នុយក្លេអូទីតទាំងនេះរលួយយ៉ាងឆាប់រហ័ស។ អាយុកាលរបស់ tRNA នៅក្នុងកោសិកាមនុស្សមានចាប់ពីប៉ុន្មាននាទីទៅច្រើនម៉ោង។ ប្រភេទខ្លះអាចមានរយៈពេលរហូតដល់មួយថ្ងៃ។ ហើយប្រសិនបើយើងនិយាយអំពី nucleotides ដូចគ្នានៅក្នុងបាក់តេរីនោះ ពេលវេលាគឺខ្លីជាង - រហូតដល់ច្រើនម៉ោង។ លើសពីនេះ រចនាសម្ព័ន្ធ និងមុខងាររបស់ tRNA គឺស្មុគស្មាញពេកសម្រាប់ម៉ូលេគុលដើម្បីក្លាយជាធាតុចម្បងនៃជីវមណ្ឌលរបស់ផែនដី។
tRNAs ទាំងអស់មានលក្ខណៈទូទៅទាំងនៅក្នុងរចនាសម្ព័ន្ធចម្បងរបស់ពួកគេ និងតាមរបៀបដែលខ្សែសង្វាក់ polynucleotide ត្រូវបានបត់ចូលទៅក្នុងរចនាសម្ព័ន្ធបន្ទាប់បន្សំ ដោយសារអន្តរកម្មរវាងមូលដ្ឋាននៃសំណល់នុយក្លេអូទីត។
រចនាសម្ព័ន្ធបឋមនៃ tRNA
tRNAs គឺជាម៉ូលេគុលតូចមួយដែលប្រវែងនៃខ្សែសង្វាក់របស់វាប្រែប្រួលពី 74 ទៅ 95 សំណល់នុយក្លេអូទីត។ tRNA ទាំងអស់មានចុង 3 អ៊ីងដូចគ្នា បង្កើតចេញពីសំណល់ស៊ីតូស៊ីនពីរ និងសំណល់អាឌីណូស៊ីនមួយ (ចុង CCA) ។ វាគឺជាអាឌីណូស៊ីនស្ថានីយ 3 អ៊ីញដែលភ្ជាប់ទៅនឹងសំណល់អាស៊ីតអាមីណូអំឡុងពេលបង្កើត aminoacyl-tRNA ។ ចុង CCA ត្រូវបានភ្ជាប់ទៅនឹង tRNAs ជាច្រើនដោយអង់ស៊ីមពិសេស។ នុយក្លេអូទីត triplet បំពេញបន្ថែមទៅ codon សម្រាប់អាស៊ីតអាមីណូ (anticodon) មានទីតាំងនៅកណ្តាលខ្សែសង្វាក់ tRNA ។ នៅទីតាំងជាក់លាក់នៅក្នុងលំដាប់ ស្ទើរតែគ្រប់ប្រភេទនៃ tRNA មានសំណល់នុយក្លេអូទីតដូចគ្នា (រក្សាទុក)។ មុខតំណែងមួយចំនួនអាចមានទាំង purine ឬមូលដ្ឋាន pyrimidine តែប៉ុណ្ណោះ (ពួកវាត្រូវបានគេហៅថាសំណល់ semiconservative) ។
ម៉ូលេគុល tRNA ទាំងអស់ត្រូវបានកំណត់លក្ខណៈដោយវត្តមាននៃចំនួនដ៏ច្រើន (រហូតដល់ 25% នៃសំណល់ទាំងអស់) នៃ nucleosides ដែលត្រូវបានកែប្រែផ្សេងៗ ដែលជារឿយៗត្រូវបានគេហៅថាអនីតិជន។ ពួកវាត្រូវបានបង្កើតឡើងនៅកន្លែងផ្សេងៗនៃម៉ូលេគុល ក្នុងករណីជាច្រើនត្រូវបានកំណត់យ៉ាងល្អ ជាលទ្ធផលនៃការកែប្រែសំណល់នុយក្លេអូស៊ីតធម្មតាដោយអង់ស៊ីមពិសេស។
រចនាសម្ព័ន្ធបន្ទាប់បន្សំនៃ tRNA
ការបត់ខ្សែសង្វាក់ចូលទៅក្នុងរចនាសម្ព័ន្ធបន្ទាប់បន្សំកើតឡើងដោយសារតែការបំពេញគ្នាទៅវិញទៅមកនៃផ្នែកខ្សែសង្វាក់។ បំណែកខ្សែសង្វាក់ទាំងបីក្លាយជាការបំពេញបន្ថែមនៅពេលបត់លើខ្លួនវា បង្កើតជារចនាសម្ព័ន្ធដូចម្ជុលសក់។ លើសពីនេះទៀតចុងបញ្ចប់ 5 "គឺបំពេញបន្ថែមទៅតំបន់ដែលនៅជិតចុង 3" នៃខ្សែសង្វាក់ជាមួយនឹងការរៀបចំប្រឆាំងនឹងប៉ារ៉ាឡែលរបស់ពួកគេ; ពួកគេបង្កើតអ្វីដែលគេហៅថា ដើមអ្នកទទួល។ លទ្ធផលគឺជារចនាសម្ព័ន្ធដែលកំណត់ដោយវត្តមាននៃដើមចំនួនបួននិងរង្វិលជុំចំនួនបីដែលត្រូវបានគេហៅថា "ស្លឹក clover" ។ ដើមនិងរង្វិលជុំបង្កើតជាសាខា។ នៅផ្នែកខាងក្រោមគឺជាសាខា anticodon ដែលផ្ទុក anticodon triplet ជាផ្នែកនៃរង្វិលជុំរបស់វា។ នៅខាងឆ្វេងនិងខាងស្តាំនៃនេះគឺជាសាខា D និង T ដែលត្រូវបានដាក់ឈ្មោះរៀងគ្នាសម្រាប់វត្តមាននៃ nucleosides ដែលត្រូវបានអភិរក្សមិនធម្មតា dihydrouridine (D) និង thymidine (T) នៅក្នុងរង្វិលជុំរបស់ពួកគេ។ លំដាប់នុយក្លេអូទីតនៃ tRNA ទាំងអស់ដែលបានសិក្សាអាចបត់ចូលទៅក្នុងរចនាសម្ព័ន្ធស្រដៀងគ្នា។ បន្ថែមពីលើរង្វិលជុំ cloverleaf ទាំងបី tRNA ក៏មានរង្វិលជុំបន្ថែម ឬអថេរ (V-loop) ផងដែរ។ ទំហំរបស់វាប្រែប្រួលយ៉ាងខ្លាំងក្នុងចំណោម tRNAs ផ្សេងៗគ្នា ប្រែប្រួលពី 4 ទៅ 21 nucleotides ហើយយោងទៅតាមទិន្នន័យចុងក្រោយបំផុត រហូតដល់ទៅ 24 nucleotides ។
រចនាសម្ព័នធរណីមាត្រនៃ tRNA
ដោយសារតែអន្តរកម្មនៃធាតុនៃរចនាសម្ព័ន្ធបន្ទាប់បន្សំរចនាសម្ព័ន្ធទីបីត្រូវបានបង្កើតឡើងដែលត្រូវបានគេហៅថាទម្រង់ L ដោយសារតែវាស្រដៀងនឹងអក្សរឡាតាំង L (រូបភាព 2 និង 3) ។ ដោយការជង់មូលដ្ឋាន ដើមអ្នកទទួល និងដើម cloverleaf T បង្កើតជា helix ពីរជាប់គ្នា ហើយដើមពីរទៀតគឺ anticodon និង D បង្កើតជា helix ពីរបន្តបន្ទាប់ទៀត។ ក្នុងករណីនេះ D- និង T-loops ត្រូវបាននាំមកជិតគ្នា ហើយត្រូវបានភ្ជាប់ជាមួយគ្នាដោយការបង្កើតគូមូលដ្ឋានបន្ថែម ដែលជាញឹកញាប់មិនធម្មតា។ សំណល់អភិរក្សឬពាក់កណ្តាលអភិរក្សជាក្បួនចូលរួមក្នុងការបង្កើតគូទាំងនេះ។ អន្តរកម្មទីបីស្រដៀងគ្នានេះ ភ្ជាប់ផ្នែកខ្លះទៀតនៃរចនាសម្ព័ន្ធ L
រីបូសូម RNA
អាស៊ីត Ribosomal ribonucleic (rRNA) គឺជាម៉ូលេគុល RNA ជាច្រើនដែលបង្កើតជាមូលដ្ឋាននៃ ribosome ។ មុខងារចម្បងរបស់ rRNA គឺដើម្បីអនុវត្តដំណើរការបកប្រែ - ការអានព័ត៌មានពី mRNA ដោយប្រើម៉ូលេគុលអាដាប់ទ័រ tRNA និងជំរុញការបង្កើតចំណង peptide រវាងអាស៊ីតអាមីណូដែលភ្ជាប់ទៅនឹង tRNA ។ Ribosomal RNA បង្កើតបានប្រហែល 80% នៃ RNA សរុបរបស់កោសិកាមួយ។ វាត្រូវបានអ៊ិនកូដដោយហ្សែនដែលរកឃើញនៅលើ DNA នៃក្រូម៉ូសូមជាច្រើនដែលមានទីតាំងនៅតំបន់មួយនៃ nucleolus ដែលត្រូវបានគេស្គាល់ថាជាអ្នករៀបចំ nucleolar ។
លំដាប់នៃមូលដ្ឋាននៅក្នុង rRNA គឺស្រដៀងគ្នានៅក្នុងសារពាង្គកាយទាំងអស់ ពីបាក់តេរីទៅសត្វ។ rRNA ត្រូវបានរកឃើញនៅក្នុង cytoplasm ដែលវាភ្ជាប់ទៅនឹងម៉ូលេគុលប្រូតេអ៊ីន បង្កើតជាកោសិកាកោសិកាដែលហៅថា ribosomes។ ការសំយោគប្រូតេអ៊ីនកើតឡើងនៅលើ ribosomes ។ នៅទីនេះ "កូដ" ដែលមាននៅក្នុង mRNA ត្រូវបានបកប្រែទៅជាលំដាប់អាស៊ីតអាមីណូនៃខ្សែសង្វាក់ polypeptide ។
ផ្ទេរ RNA
ផ្ទេរ RNA, tRNA, គឺជាអាស៊ីត ribonucleic ដែលមានមុខងារដឹកជញ្ជូនអាស៊ីតអាមីណូទៅកាន់កន្លែងសំយោគប្រូតេអ៊ីន។ tRNAs ក៏ចូលរួមដោយផ្ទាល់ក្នុងការពង្រីកខ្សែសង្វាក់ polypeptide ដោយការចូលរួម - ស្មុគ្រស្មាញជាមួយអាស៊ីតអាមីណូ - ទៅនឹង mRNA codon និងផ្តល់នូវការអនុលោមភាពស្មុគស្មាញដែលចាំបាច់សម្រាប់ការបង្កើតចំណង peptide ថ្មី។
អាស៊ីតអាមីណូនីមួយៗមាន tRNA ផ្ទាល់ខ្លួន។
tRNA គឺជា RNA តែមួយខ្សែ ប៉ុន្តែនៅក្នុងទម្រង់មុខងាររបស់វា វាមានការអនុលោមតាម cloverleaf ។ វាមានបួនផ្នែកសំខាន់ៗដែលបំពេញមុខងារផ្សេងៗគ្នា។ ឧបករណ៍ទទួល "ដើម" ត្រូវបានបង្កើតឡើងដោយផ្នែកស្ថានីយដែលភ្ជាប់គ្នាពីរនៃ tRNA ។ វាមានគូមូលដ្ឋានចំនួនប្រាំពីរ។ ចុង 3" នៃដើមនេះវែងជាងបន្តិច ហើយបង្កើតបានជាតំបន់ដែលមានខ្សែតែមួយដែលបញ្ចប់ដោយលំដាប់ CCA ជាមួយនឹងក្រុម OH ឥតគិតថ្លៃ។ អាស៊ីតអាមីណូដែលបានដឹកជញ្ជូនត្រូវបានភ្ជាប់ទៅចុងនេះ។ សាខាទាំងបីដែលនៅសល់គឺជាលំដាប់នៃ nucleotide ផ្គូផ្គងបំពេញបន្ថែមដែលបញ្ចប់។ នៅក្នុងតំបន់ដែលមិនផ្គូផ្គងដែលបង្កើតជារង្វិលជុំ។ ផ្នែកកណ្តាលនៃសាខាទាំងនេះ - anticodon - មាន nucleotides ប្រាំគូ និងមាន anticodon នៅកណ្តាលរង្វិលជុំរបស់វា។ anticodon គឺជា nucleotides បីដែលបំពេញបន្ថែមទៅនឹង mRNA codon ដែលបំប្លែងអាស៊ីតអាមីណូដែលបានដឹកជញ្ជូន។ ដោយ tRNA នេះទៅកន្លែងនៃការសំយោគ peptide ។
រវាងអ្នកទទួល និង អង់ទីកូដុន មានសាខាពីរចំហៀង។ នៅក្នុងរង្វិលជុំពួកវាមានមូលដ្ឋានដែលបានកែប្រែ -dihydrouridine (D-loop) និង T?C triplet នៅឯណា? - pseudorain (T?C-loop) ។ រវាងសាខា aiticodon និង T?C មានរង្វិលជុំបន្ថែម រួមទាំងពី 3-5 ទៅ 13-21 nucleotides ។
អាស៊ីតអាមីណូត្រូវបានភ្ជាប់ជាកូវ៉ាឡង់ទៅចុង 3 "នៃម៉ូលេគុលដោយប្រើអង់ស៊ីម aminoacyl-tRNA synthetase ដែលជាក់លាក់សម្រាប់ប្រភេទនីមួយៗនៃ tRNA ។
tRNA បម្រើជាម៉ូលេគុលមធ្យមរវាង codon triplet ក្នុង mRNA និងលំដាប់អាស៊ីតអាមីណូនៃខ្សែសង្វាក់ polypeptide ។ tRNA មានប្រហែល 15% នៃ RNA កោសិកាទាំងអស់; RNA ទាំងនេះមានខ្សែសង្វាក់ polynucleotide ខ្លីបំផុត - វាមានជាមធ្យម 80 nucleotides ។ កោសិកានីមួយៗមានម៉ូលេគុល tRNA ច្រើនជាង 20 ផ្សេងគ្នា។ ម៉ូលេគុល tRNA ទាំងអស់មានរចនាសម្ព័ន្ធមូលដ្ឋានស្រដៀងគ្នា។ នៅចុង 5' នៃម៉ូលេគុល tRNA តែងតែមាន guanine ហើយនៅចុង 3' មានលំដាប់នៃមូលដ្ឋាន CCA ។
លំដាប់នុយក្លេអូទីតនៅក្នុងម៉ូលេគុលដែលនៅសល់ប្រែប្រួល ហើយអាចមានមូលដ្ឋាន "មិនធម្មតា" ដូចជា inosine និង pseudouracil ។
លំដាប់នៃមូលដ្ឋាននៅក្នុង anticodon triplet ត្រូវគ្នាយ៉ាងតឹងរឹងទៅនឹងអាស៊ីតអាមីណូដែលម៉ូលេគុល tRNA នេះផ្ទុក។
អង្ករ។ ៣.
អាស៊ីតអាមីណូនីមួយៗត្រូវបានភ្ជាប់ទៅនឹង tRNAs ជាក់លាក់មួយរបស់វា ដោយមានការចូលរួមពីអង់ស៊ីម aminoacyl-tRNA synthase ។ នេះបណ្តាលឱ្យមានការបង្កើតស្មុគស្មាញ animoacid-tRNA ដែលត្រូវបានគេស្គាល់ថា animoacyl-tRNA ដែលថាមពលចំណងរវាងស្ថានីយ A nucleotide ក្នុង CCA triplet និងអាស៊ីតអាមីណូគឺគ្រប់គ្រាន់ដើម្បីអនុញ្ញាតឱ្យមានទំនាក់ទំនងជាបន្តបន្ទាប់ជាមួយអាស៊ីតអាមីណូដែលនៅជិតខាង។ ដូច្នេះខ្សែសង្វាក់ polypeptide ត្រូវបានសំយោគ។
លក្ខណៈពិសេសមួយនៃ tRNA គឺវត្តមាននៃមូលដ្ឋានមិនធម្មតានៅក្នុងវាដែលកើតឡើងជាលទ្ធផលនៃការកែប្រែគីមីបន្ទាប់ពីការរួមបញ្ចូលនៃមូលដ្ឋានធម្មតានៅក្នុងខ្សែសង្វាក់ polynucleotide ។ មូលដ្ឋានដែលបានផ្លាស់ប្តូរទាំងនេះកំណត់ពីភាពចម្រុះនៃរចនាសម្ព័ន្ធដ៏អស្ចារ្យនៃ tRNAs នៅក្នុងផែនការទូទៅនៃរចនាសម្ព័ន្ធរបស់ពួកគេ។ ការចាប់អារម្មណ៍ខ្លាំងបំផុតគឺការកែប្រែមូលដ្ឋានបង្កើត anticodon ដែលប៉ះពាល់ដល់ភាពជាក់លាក់នៃអន្តរកម្មរបស់វាជាមួយ codon ។ ឧទាហរណ៍ inosine មូលដ្ឋាន atypical ដែលជួនកាលត្រូវបានគេរកឃើញនៅក្នុងទីតាំងទី 1 នៃ tRNA anticodon មានសមត្ថភាពរួមបញ្ចូលគ្នាជាមួយនឹងមូលដ្ឋានទីបីផ្សេងគ្នានៃ mRNA codon - U, C និង A. ចាប់តាំងពីលក្ខណៈពិសេសមួយនៃកូដហ្សែនគឺ degeneracy របស់វា អាស៊ីតអាមីណូជាច្រើនត្រូវបានអ៊ិនគ្រីបដោយ codons ជាច្រើន ដែលជាធម្មតាមានភាពខុសគ្នានៅក្នុងមូលដ្ឋានទីបីរបស់វា។ ដោយសារតែការចងមិនជាក់លាក់នៃមូលដ្ឋាន anticodon ដែលបានកែប្រែ tRNA មួយទទួលស្គាល់ codons មានន័យដូចមួយចំនួន។