Struktura drugorzędowa RNA. Struktura i funkcja kwasów rybonukleinowych
Cząsteczka RNA jest również polimerem, którego monomerami są rybonukleotydy; RNA jest cząsteczką jednoniciową. Jest zbudowany w taki sam sposób, jak jedna z nici DNA. Nukleotydy RNA są podobne do nukleotydów DNA, chociaż nie są z nimi identyczne. Jest ich również cztery i składają się z reszt zasad azotowych, pentozy i kwasu fosforowego. Trzy zasady azotowe są dokładnie takie same jak w DNA: A, G I C. Jednak zamiast T DNA w RNA zawiera zasadę pirymidynową o podobnej budowie – uracyl ( U). Główną różnicą między DNA i RNA jest natura węglowodanów: w nukleotydach DNA monosacharydem jest deoksyryboza, a w RNA ryboza. Połączenie między nukleotydami odbywa się, podobnie jak w DNA, poprzez resztę cukru i kwasu fosforowego. W przeciwieństwie do DNA, którego zawartość w komórkach niektórych organizmów jest stała, zawartość RNA w nich jest zmienna. Jest on zauważalnie wyższy tam, gdzie zachodzi intensywna synteza.
Ze względu na funkcje jakie pełnią wyróżnia się kilka typów RNA.
Przenieść RNA (tRNA). Cząsteczki tRNA są najkrótsze: składają się z zaledwie 80-100 nukleotydów. Masa cząsteczkowa takich cząstek wynosi 25-30 tys. Transferowe RNA zawarte są głównie w cytoplazmie komórki. Ich funkcją jest przenoszenie aminokwasów do rybosomów, do miejsca syntezy białek. Z całkowitej zawartości RNA w komórkach tRNA stanowi około 10%.
Rybosomalny RNA (rRNA). Są to duże cząsteczki: zawierają odpowiednio 3-5 tysięcy nukleotydów, ich masa cząsteczkowa sięga 1-1,5 miliona, rybosomalne RNA stanowią znaczną część rybosomu. Z całkowitej zawartości RNA w komórce rRNA stanowi około 90%.
Komunikator RNA (mRNA) lub informacyjny RNA (mRNA) znajduje się w jądrze i cytoplazmie. Jego funkcją jest przekazywanie informacji o strukturze białka z DNA do miejsca syntezy białka w rybosomach. mRNA stanowi około 0,5-1% całkowitej zawartości RNA w komórce. Rozmiar mRNA jest bardzo zróżnicowany - od 100 do 10 000 nukleotydów.
Wszystkie typy RNA są syntetyzowane na DNA, które służy jako rodzaj matrycy.
DNA jest nośnikiem informacji dziedzicznej.
Każde białko jest reprezentowane przez jeden lub więcej łańcuchów polipeptydowych. Nazywa się odcinek DNA, który niesie informację o jednym łańcuchu polipeptydowym genom. Całość cząsteczek DNA w komórce pełni rolę nośnika informacji genetycznej. Informacja genetyczna przekazywana jest zarówno z komórek macierzystych do komórek potomnych, jak i od rodziców do dzieci. Gen jest jednostką genetyczną, Lub informacje dziedziczne.
DNA jest nośnikiem informacji genetycznej w komórce – nie uczestniczy bezpośrednio w syntezie białek. W komórkach eukariotycznych cząsteczki DNA znajdują się w chromosomach jądra i są oddzielone otoczką jądrową od cytoplazmy, gdzie zachodzi synteza białek. Posłaniec przenoszący informacje jest wysyłany z jądra do rybosomów, miejsca składania białek, i może przechodzić przez pory błony jądrowej. Tym przekaźnikiem jest informacyjny RNA (mRNA). Zgodnie z zasadą komplementarności syntetyzowany jest na DNA przy udziale enzymu zwanego RNA polimeraza.

Informacyjny RNA jest cząsteczką jednoniciową, a transkrypcja zachodzi z jednej nici dwuniciowej cząsteczki DNA. Nie jest to kopia całej cząsteczki DNA, a jedynie jej część – jeden gen u eukariontów lub grupa sąsiadujących ze sobą genów, które niosą informację o strukturze białek niezbędnych do pełnienia jednej funkcji u prokariotów. Ta grupa genów nazywa się operon. Na początku każdego operonu znajduje się swego rodzaju lądowisko dla polimerazy RNA, tzw promotor.jest to specyficzna sekwencja nukleotydów DNA, którą enzym „rozpoznaje” ze względu na powinowactwo chemiczne. Tylko poprzez przyłączenie się do promotora polimeraza RNA może rozpocząć syntezę RNA. Po dotarciu do końca operonu enzym napotyka sygnał (w postaci określonej sekwencji nukleotydów) wskazujący koniec odczytu. Gotowy mRNA opuszcza DNA i trafia do miejsca syntezy białka.
W procesie transkrypcji można wyróżnić cztery etapy: 1) Wiązanie RNA-polimeraza z promotorem; 2) inicjacja– początek syntezy. Polega na utworzeniu pierwszego wiązania fosfodiestrowego pomiędzy ATP lub GTP a drugim nukleotydem syntetyzowanej cząsteczki RNA; 3) wydłużenie– wzrost łańcucha RNA; te. sekwencyjne dodawanie nukleotydów do siebie w kolejności, w jakiej ich komplementarne nukleotydy pojawiają się w transkrybowanej nici DNA. Szybkość wydłużania wynosi 50 nukleotydów na sekundę; 4) zakończenie– zakończenie syntezy RNA.
Po przejściu przez pory błony jądrowej mRNA zostaje wysłany do rybosomów, gdzie zostaje rozszyfrowana informacja genetyczna - przetłumaczona z „języka” nukleotydów na „język” aminokwasów. Synteza łańcuchów polipeptydowych z wykorzystaniem matrycy mRNA, która zachodzi w rybosomach, nazywa się audycja(Tłumaczenie łacińskie - tłumaczenie).
Aminokwasy, z których syntetyzowane są białka, dostarczane są do rybosomów za pomocą specjalnych RNA zwanych transferowymi RNA (tRNA). W komórce jest tyle różnych tRNA, ile jest kodonów kodujących aminokwasy. Na górze „liście” każdego tRNA znajduje się sekwencja trzech nukleotydów, które są komplementarne do nukleotydów kodonu w mRNA. Dzwonią do niej antykodon. Specjalny enzym, kodaza, rozpoznaje tRNA i przyłącza aminokwas do „ogonka liścia” – tylko ten kodowany przez triplet komplementarny do antykodonu. Utworzenie wiązania kowalencyjnego pomiędzy tRNA i jego „własnym” aminokwasem wymaga energii jednej cząsteczki ATP.
 Aby aminokwas mógł zostać włączony do łańcucha polipeptydowego, musi odłączyć się od tRNA. Staje się to możliwe, gdy tRNA dostanie się do rybosomu, a antykodon rozpoznaje swój kodon w mRNA. Rybosom ma dwa miejsca do wiązania dwóch cząsteczek tRNA. W jednym z tych obszarów, tzw akceptor tRNA przybywa z aminokwasem i przyłącza się do jego kodonu (I). Czy ten aminokwas przyłącza się do siebie (akceptuje) rosnący łańcuch białkowy (II)? Tworzy się między nimi wiązanie peptydowe. tRNA, który jest teraz przyłączony wraz z kodonem mRNA in dawca część rybosomu. Nowy tRNA pojawia się w pustym miejscu akceptorowym, związany z aminokwasem, który jest szyfrowany przez następny kodon (III). Odłączony łańcuch polipeptydowy jest tutaj ponownie przenoszony z miejsca dawcy i przedłużany o jeszcze jedno ogniwo. Aminokwasy w rosnącym łańcuchu są połączone w takiej kolejności, w jakiej kodony je kodony są zlokalizowane w mRNA.
Aby aminokwas mógł zostać włączony do łańcucha polipeptydowego, musi odłączyć się od tRNA. Staje się to możliwe, gdy tRNA dostanie się do rybosomu, a antykodon rozpoznaje swój kodon w mRNA. Rybosom ma dwa miejsca do wiązania dwóch cząsteczek tRNA. W jednym z tych obszarów, tzw akceptor tRNA przybywa z aminokwasem i przyłącza się do jego kodonu (I). Czy ten aminokwas przyłącza się do siebie (akceptuje) rosnący łańcuch białkowy (II)? Tworzy się między nimi wiązanie peptydowe. tRNA, który jest teraz przyłączony wraz z kodonem mRNA in dawca część rybosomu. Nowy tRNA pojawia się w pustym miejscu akceptorowym, związany z aminokwasem, który jest szyfrowany przez następny kodon (III). Odłączony łańcuch polipeptydowy jest tutaj ponownie przenoszony z miejsca dawcy i przedłużany o jeszcze jedno ogniwo. Aminokwasy w rosnącym łańcuchu są połączone w takiej kolejności, w jakiej kodony je kodony są zlokalizowane w mRNA.
Kiedy na rybosomie pojawi się jedna z trzech trójek ( UAA, UAG, UGA), które są „znakami interpunkcyjnymi” pomiędzy genami, żadne tRNA nie może zająć miejsca w miejscu akceptorowym. Faktem jest, że nie ma antykodonów komplementarnych do sekwencji nukleotydowych „znaków interpunkcyjnych”. Odłączona nić nie ma się do czego przyczepić w miejscu akceptorowym i opuszcza rybosom. Synteza białek została zakończona.
U prokariotów synteza białek rozpoczyna się od kodonu SIERPIEŃ, znajdujący się na pierwszym miejscu w kopii każdego genu, zajmuje w rybosomie taką pozycję, z którą oddziałuje antykodon specjalnego tRNA z nim połączonego formylomentionina. Ta zmodyfikowana forma aminokwasu metioniny natychmiast wchodzi do miejsca dawcy i pełni funkcję dużej litery w zdaniu - synteza dowolnego łańcucha polipeptydowego rozpoczyna się od niej w komórce bakteryjnej. Kiedy trójka SIERPIEŃ nie znajduje się na pierwszym miejscu, ale wewnątrz kopii genu; koduje aminokwas metioninę. Po zakończeniu syntezy łańcucha polipeptydowego formylometionina zostaje od niego odszczepiona i nie występuje w gotowym białku.
 Aby zwiększyć produkcję białka, mRNA często przechodzi nie przez jeden, ale przez kilka rybosomów jednocześnie. Struktura ta, połączona przez jedną cząsteczkę mRNA, nazywa się polisom. Każdy rybosom w tym przypominającym kulkę przenośniku taśmowym syntetyzuje te same białka.
Aby zwiększyć produkcję białka, mRNA często przechodzi nie przez jeden, ale przez kilka rybosomów jednocześnie. Struktura ta, połączona przez jedną cząsteczkę mRNA, nazywa się polisom. Każdy rybosom w tym przypominającym kulkę przenośniku taśmowym syntetyzuje te same białka.
Aminokwasy są w sposób ciągły dostarczane do rybosomów za pomocą tRNA. Po oddaniu aminokwasu tRNA opuszcza rybosom i łączy się za pomocą kodazy. Wysoka spójność wszystkich „usług rośliny” do produkcji białek umożliwia syntezę łańcuchów polipeptydowych składających się z setek aminokwasów w ciągu kilku sekund.
Właściwości kodu genetycznego. Dzięki procesowi transkrypcji w komórce informacja przekazywana jest z DNA na białko
DNA → mRNA → białko
Informacja genetyczna zawarta w DNA i mRNA zawarta jest w sekwencji nukleotydów w cząsteczkach.
 W jaki sposób informacja jest przekazywana z „języka” nukleotydów do „języka” aminokwasów? Tłumaczenie to odbywa się przy użyciu kodu genetycznego. Kod lub szyfr, to system symboli służący do tłumaczenia jednej formy informacji na inną. Kod genetyczny to system zapisu informacji o sekwencji aminokwasów w białkach na podstawie sekwencji nukleotydów w mRNA.
W jaki sposób informacja jest przekazywana z „języka” nukleotydów do „języka” aminokwasów? Tłumaczenie to odbywa się przy użyciu kodu genetycznego. Kod lub szyfr, to system symboli służący do tłumaczenia jednej formy informacji na inną. Kod genetyczny to system zapisu informacji o sekwencji aminokwasów w białkach na podstawie sekwencji nukleotydów w mRNA.
Jakie właściwości ma kod genetyczny?
Kod jest potrójny. RNA zawiera cztery nukleotydy: A, G, C, U. Gdybyśmy próbowali oznaczyć jeden aminokwas jednym nukleotydem, wówczas 16 z 20 aminokwasów pozostałoby niekodowanych. Dwuliterowy kod szyfrowałby 16 aminokwasów. Natura stworzyła trzyliterowy kod. To znaczy, że Każdy z 20 aminokwasów jest kodowany przez sekwencję trzech nukleotydów zwaną tripletem lub kodonem.
Kod jest zdegenerowany. To znaczy, że Każdy aminokwas jest kodowany przez więcej niż jeden kodon. Wyjątki: meteonina i tryptofan, z których każdy jest kodowany przez jedną trójkę.
Kod jest jasny. Każdy kodon koduje tylko jeden aminokwas.
Pomiędzy genami znajdują się „znaki interpunkcyjne”. W tekście drukowanym na końcu każdego wyrażenia znajduje się kropka. Akapit składa się z kilku powiązanych ze sobą wyrażeń. W języku informacji genetycznej takim akapitem jest operon i komplementarny do niego mRNA. Każdy gen w operonie prokariotycznym lub oddzielny gen eukariotyczny koduje jeden łańcuch polipeptydowy – frazę. Ponieważ w niektórych przypadkach z matrycy mRNA powstaje sekwencyjnie kilka różnych łańcuchów polipeptydowych, należy je od siebie oddzielić. W tym celu w roku genetycznym wyróżnia się trzy specjalne trojaczki – UAA, UAG, UGA, z których każda wskazuje na ustanie syntezy jednego łańcucha polipeptydowego. Zatem te trójki pełnią funkcję znaków interpunkcyjnych. Znajdują się na końcu każdego genu.
W genie nie ma „znaków interpunkcyjnych”.
Kod jest uniwersalny. Kod genetyczny jest taki sam dla wszystkich stworzeń żyjących na Ziemi. U bakterii i grzybów, pszenicy i bawełny, ryb i robaków, żab i ludzi te same trojaczki kodują te same aminokwasy.

Zasady replikacji DNA. Proces ten zapewnia ciągłość materiału genetycznego w pokoleniach komórek i organizmów replikacja - podwojenie cząsteczek DNA. W tym złożonym procesie bierze udział kompleks kilku enzymów i białek, które nie mają aktywności katalitycznej, a które są niezbędne do nadania łańcuchom polinukleotydowym pożądanej konformacji. W wyniku replikacji powstają dwie identyczne podwójne helisy DNA. Te tak zwane cząsteczki potomne nie różnią się od siebie ani od pierwotnej cząsteczki macierzystego DNA. Replikacja zachodzi w komórce przed podziałem, więc każda komórka potomna otrzymuje dokładnie te same cząsteczki DNA, które miała komórka macierzysta. Proces replikacji opiera się na kilku zasadach:
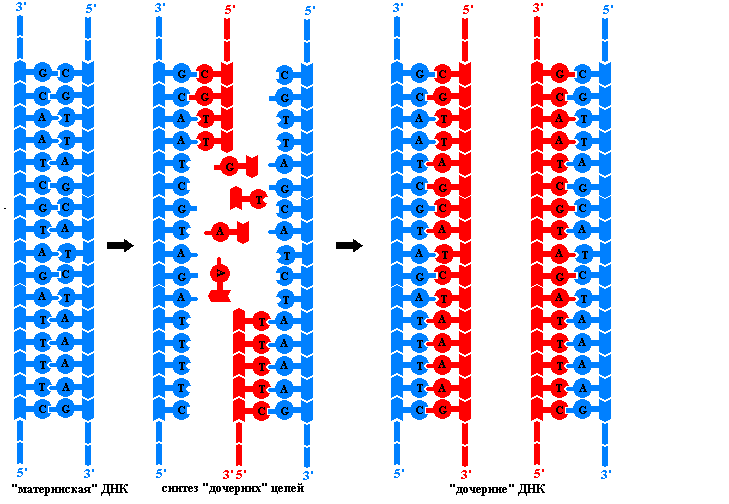
Tylko w tym przypadku polimerazy DNA są w stanie przemieszczać się wzdłuż nici macierzystych i wykorzystywać je jako matryce do bezbłędnej syntezy nici potomnych. Jednak całkowite rozwinięcie helis składających się z wielu milionów par nukleotydów wiąże się z tak znaczną liczbą obrotów i takimi kosztami energii, które są niemożliwe w warunkach komórkowych. Dlatego replikacja u eukariontów rozpoczyna się jednocześnie w niektórych miejscach cząsteczki DNA. Obszar pomiędzy dwoma punktami, w którym rozpoczyna się synteza łańcuchów potomnych, nazywa się replikon. On jest jednostka replikacji.

Każda cząsteczka DNA komórki eukariotycznej zawiera wiele replikonów. W każdym replikonie widać widełki replikacyjne – tę część cząsteczki DNA, która uległa już rozplątaniu pod wpływem specjalnych enzymów. Każda nić w widelcu służy jako szablon do syntezy komplementarnej nici potomnej. Podczas replikacji widełki poruszają się wzdłuż cząsteczki macierzystej i rozwijają się nowe odcinki DNA. Ponieważ polimerazy DNA mogą poruszać się tylko w jednym kierunku wzdłuż nici matrycy, a nici są zorientowane antyrównolegle, w każdym widelcu syntetyzowane są jednocześnie dwa różne kompleksy enzymatyczne. Co więcej, w każdym widelcu jeden łańcuch potomny (wiodący) rośnie w sposób ciągły, podczas gdy drugi (opóźniony) łańcuch jest syntetyzowany w oddzielnych fragmentach o długości kilku nukleotydów. Takie enzymy, nazwane na cześć japońskiego naukowca, który je odkrył fragmenty Okazaki, są usieciowane ligazą DNA, tworząc ciągły łańcuch. Mechanizm tworzenia nici potomnych DNA przez fragmenty nazywa się nieciągłym.
Wymóg starterowania polimerazy DNA nie jest w stanie zainicjować syntezy nici wiodącej, ani syntezy fragmentów nici opóźnionej Okazaki. Może jedynie wydłużyć istniejącą nić polinukleotydową poprzez kolejne dodanie deoksyrybonukleotydów do jej końca 3’-OH. Skąd pochodzi początkowy region 5'-końcowy rosnącego łańcucha DNA? Jest syntetyzowany na matrycy DNA przez specjalną polimerazę RNA zwaną prymas(Podkład języka angielskiego - materiał siewny). Rozmiar startera rybonukleotydowego jest niewielki (mniej niż 20 nukleotydów) w porównaniu z rozmiarem łańcucha DNA utworzonego przez poimerazę DNA. Skończywszy ją Funkcjonować Starter RNA jest usuwany przez specjalny enzym, a powstała w tym procesie szczelina jest zamykana przez polimerazę DNA, która jako starter wykorzystuje koniec 3'-OH sąsiadującego fragmentu Okazaki.
Problem niedostatecznej replikacji końców liniowych cząsteczek DNA. Usuwanie skrajnych starterów RNA, komplementarne do końców 3' obu nici liniowej macierzystej cząsteczki DNA, prowadzi do tego, że nici potomne są krótsze niż 10-20 nukleotydów. Jest to problem niedostatecznej replikacji końców cząsteczek liniowych.
Problem niedostatecznej replikacji końców 3' liniowych cząsteczek DNA rozwiązują komórki eukariotyczne za pomocą specjalnego enzymu - telomeraza.
Telomeraza jest polimerazą DNA, która uzupełnia 3'-końcowe cząsteczki DNA chromosomów o krótkie powtarzające się sekwencje. Umieszczone jeden za drugim tworzą regularną strukturę końcową o długości do 10 tysięcy nukleotydów. Oprócz części białkowej telomeraza zawiera RNA, który działa jako matryca do wydłużania powtórzeń DNA.
Schemat wydłużania końców cząsteczek DNA. Najpierw następuje komplementarne wiązanie wystającego końca DNA z regionem matrycowym RNA telomerazy, następnie telomeraza wydłuża DNA wykorzystując jego koniec 3’-OH jako starter i RNA zawarty w enzymie jako matrycę. Ten etap nazywa się elongacją. Następnie następuje translokacja, tj. ruch DNA wydłużony o jedno powtórzenie w stosunku do enzymu. Następnie następuje wydłużenie i kolejna translokacja.
W rezultacie powstają wyspecjalizowane struktury końcowe chromosomów. Składają się z wielokrotnie powtarzających się krótkich sekwencji DNA i specyficznych białek.
Synteza prekursorów rRNA i tRNA przebiega podobnie do syntezy ire-mRNA. Pierwotny transkrypt rybosomalnego RNA nie zawiera intronów i pod działaniem specyficznych RNaz ulega rozszczepieniu, tworząc 28S-, 18S- i 5.8S-rRNA; Synteza 5S-pRNA odbywa się przy udziale polimerazy RNA III.
rRNA i tRNA.
Pierwotne transkrypty tRNA są również przekształcane w formy dojrzałe w wyniku częściowej hydrolizy.
Wszystkie typy RNA biorą udział w biosyntezie białek, jednak ich funkcje w tym procesie są różne. Rolę macierzy określającej pierwszorzędową strukturę białek pełni informacyjny RNA (mRNA).W badaniu mechanizmów translacji istotne jest wykorzystanie bezkomórkowych systemów biosyntezy białek. Jeżeli homogenaty tkankowe inkubuje się z mieszaniną aminokwasów, z których co najmniej jeden jest znakowany, wówczas biosyntezę białek można wykryć poprzez włączenie znacznika do białek. Podstawowa struktura syntetyzowanego białka jest określona przez pierwotną strukturę mRNA dodanego do układu. Jeśli układ bezkomórkowy składa się z mRNA globiny (można go wyizolować z retikulocytów), syntetyzowana jest globina (łańcuchy a i 3-globiny), jeśli albumina jest syntetyzowana z mRNA albuminy wyizolowanym z hepatocytów itp.
14. Znaczenie replikacji:
a) proces ten jest ważnym mechanizmem molekularnym leżącym u podstaw wszystkich typów podziałów komórkowych u proeukariotów, b) zapewnia wszystkie rodzaje reprodukcji zarówno organizmów jednokomórkowych, jak i wielokomórkowych,
c) utrzymuje stałość komórkową
skład narządów, tkanek i organizmu w wyniku regeneracji fizjologicznej
d) zapewnia długotrwałe istnienie określonych osób;
e) zapewnia długotrwałe istnienie gatunków organizmów;
f) proces sprzyja dokładnemu podwajaniu informacji;
g) w procesie replikacji możliwe są błędy (mutacje), które mogą prowadzić do zaburzeń w syntezie białek wraz z rozwojem zmian patologicznych.
Unikalna właściwość cząsteczki DNA polegająca na powielaniu się przed podziałem komórki nazywana jest replikacją.
Szczególne właściwości natywnego DNA jako nośnika informacji dziedzicznej:
1) replikacja – tworzenie nowych łańcuchów ma charakter komplementarny;
2) samokorekta – polimeraza DNA odcina błędnie zreplikowane fragmenty (10-6);
3) naprawa – renowacja;
Procesy te zachodzą w komórce przy udziale specjalnych enzymów.
Jak działa system naprawczy Eksperymenty, które pozwoliły zidentyfikować mechanizmy regeneracji i samo istnienie tej zdolności, przeprowadzono na organizmach jednokomórkowych. Ale procesy naprawcze są nieodłącznym elementem żywych komórek zwierząt i ludzi. Niektórzy ludzie cierpią na xeroderma pigmentosum. Choroba ta jest spowodowana brakiem zdolności komórek do resyntezy uszkodzonego DNA. Xeroderma jest dziedziczna. Z czego składa się system reparacji? Cztery enzymy wspierające proces naprawy to helikaza DNA, -egzonukleaza, -polimeraza i -ligaza. Pierwszy z tych związków jest w stanie rozpoznać uszkodzenia w łańcuchu cząsteczki kwasu dezoksyrybonukleinowego. Nie tylko rozpoznaje, ale także przecina łańcuch we właściwym miejscu, aby usunąć zmodyfikowany segment cząsteczki. Sama eliminacja odbywa się za pomocą egzonukleazy DNA. Następnie z aminokwasów syntetyzuje się nową sekcję cząsteczki kwasu dezoksyrybonukleinowego, aby całkowicie zastąpić uszkodzoną sekcję. Cóż, ostatni akord tej najbardziej złożonej procedury biologicznej wykonywany jest przy użyciu enzymu ligazy DNA. Odpowiada za przyłączenie syntetyzowanego miejsca do uszkodzonej cząsteczki. Gdy wszystkie cztery enzymy wykonają swoją pracę, cząsteczka DNA zostaje całkowicie odnowiona, a wszelkie uszkodzenia należą już do przeszłości. W ten sposób mechanizmy wewnątrz żywej komórki działają harmonijnie.
Klasyfikacja Obecnie naukowcy wyróżniają następujące rodzaje systemów reparacji. Są aktywowane w zależności od różnych czynników. Należą do nich: Reaktywacja. Przywrócenie rekombinacji. Naprawa heterodupleksu. Naprawa przez wycięcie. Łączenie niehomologicznych końców cząsteczek DNA. Wszystkie organizmy jednokomórkowe mają co najmniej trzy układy enzymatyczne. Każdy z nich posiada możliwość przeprowadzenia procesu rekonwalescencji. Do systemów tych zalicza się: bezpośredni, wycinający i postreplikacyjny. Prokarioty posiadają te trzy rodzaje naprawy DNA. Jeśli chodzi o eukarionty, mają one do dyspozycji dodatkowe mechanizmy zwane Miss-mathe i Sos-repair. Biologia szczegółowo zbadała wszystkie rodzaje samoleczenia materiału genetycznego komórek.
15. Kod genetyczny to metoda kodowania sekwencji aminokwasów białek za pomocą sekwencji nukleotydów, charakterystycznej dla wszystkich organizmów żywych. Sekwencja aminokwasów w cząsteczce białka jest szyfrowana jako sekwencja nukleotydów w cząsteczce DNA i nosi nazwę kod genetyczny. Nazywa się odcinek cząsteczki DNA odpowiedzialny za syntezę jednego białka genom.
DNA wykorzystuje cztery nukleotydy - adeninę (A), guaninę (G), cytozynę (C), tyminę (T), które w literaturze rosyjskiej są oznaczone literami A, G, C i T. Litery te tworzą alfabet kod genetyczny. RNA wykorzystuje te same nukleotydy, z wyjątkiem tyminy, która jest zastąpiona podobnym nukleotydem - uracylem, który jest oznaczony literą U (U w literaturze rosyjskojęzycznej). W cząsteczkach DNA i RNA nukleotydy układają się w łańcuchy i w ten sposób uzyskuje się sekwencje liter genetycznych.
Do budowy białek w przyrodzie wykorzystuje się 20 różnych aminokwasów. Każde białko to łańcuch lub kilka łańcuchów aminokwasów w ściśle określonej kolejności. Sekwencja ta determinuje strukturę białka, a co za tym idzie, wszystkie jego właściwości biologiczne. Zestaw aminokwasów jest także uniwersalny dla niemal wszystkich organizmów żywych.
Wdrożenie informacji genetycznej w żywych komórkach (czyli synteza białka kodowanego przez gen) odbywa się za pomocą dwóch procesów macierzowych: transkrypcji (czyli syntezy mRNA na matrycy DNA) i translacji kodu genetycznego w sekwencję aminokwasową (synteza łańcucha polipeptydowego na matrycy mRNA). Trzy kolejne nukleotydy wystarczą do zakodowania 20 aminokwasów, a także sygnału stopu wskazującego koniec sekwencji białka. Zestaw trzech nukleotydów nazywany jest tripletem. Akceptowane skróty odpowiadające aminokwasom i kodonom pokazano na rysunku.
Właściwości kodu genetycznego
Triplet – znacząca jednostka kodu to kombinacja trzech nukleotydów (triplet lub kodon).
Ciągłość - pomiędzy trójkami nie ma interpunkcji, to znaczy informacja jest odczytywana w sposób ciągły.
Brak nakładania się - ten sam nukleotyd nie może być jednocześnie częścią dwóch lub więcej trypletów. (Nie dotyczy niektórych nakładających się genów w wirusach, mitochondriach i bakteriach, które kodują białka z wieloma przesunięciami ramki odczytu).
Wyjątkowość – określony kodon odpowiada tylko jednemu aminokwasowi. (Właściwość nie jest uniwersalna. Kodon UGA u Euplotes crassus koduje dwa aminokwasy – cysteinę i selenocysteinę)
Degeneracja (redundancja) - temu samemu aminokwasowi może odpowiadać kilka kodonów.
Uniwersalność – kod genetyczny działa tak samo w organizmach o różnym stopniu złożoności – od wirusów po człowieka (na tym opierają się metody inżynierii genetycznej) (Istnieje również szereg wyjątków od tej właściwości, patrz tabela w „Odmianach standardowy kod genetyczny” w tym artykule).
16.Warunki biosyntezy
Biosynteza białek wymaga informacji genetycznej z cząsteczki DNA; informacyjny RNA - nośnik tej informacji z jądra do miejsca syntezy; rybosomy - organelle, w których zachodzi sama synteza białek; zestaw aminokwasów w cytoplazmie; przenosić RNA kodujące aminokwasy i przenosić je do miejsca syntezy na rybosomach; ATP jest substancją dostarczającą energii do procesu kodowania i biosyntezy.
Gradacja
Transkrypcja- proces biosyntezy wszystkich typów RNA na matrycy DNA, który zachodzi w jądrze.
Pewna część cząsteczki DNA ulega despiracji, a wiązania wodorowe pomiędzy dwoma łańcuchami ulegają zniszczeniu pod działaniem enzymów. Na jednej nici DNA, podobnie jak na matrycy, syntetyzowana jest kopia RNA z nukleotydów zgodnie z zasadą komplementarności. W zależności od sekcji DNA, w ten sposób syntetyzowane są rybosomalne, transportowe i informacyjne RNA.
Po syntezie mRNA opuszcza jądro i kierowany jest do cytoplazmy, gdzie na rybosomach następuje synteza białek.
Audycja- proces syntezy łańcuchów polipeptydowych przeprowadzany na rybosomach, gdzie mRNA jest pośrednikiem w przekazywaniu informacji o pierwotnej strukturze białka.
Biosynteza białek składa się z szeregu reakcji.
1. Aktywacja i kodowanie aminokwasów. tRNA ma kształt liścia koniczyny, w którego centralnej pętli znajduje się antykodon tripletowy, odpowiadający kodowi konkretnego aminokwasu i kodonowi w mRNA. Każdy aminokwas jest połączony z odpowiednim tRNA za pomocą energii ATP. Tworzy się kompleks tRNA-aminokwas, który przedostaje się do rybosomów.
2. Tworzenie kompleksu mRNA-rybosom. mRNA w cytoplazmie jest połączone rybosomami na ziarnistej ER.
3. Składanie łańcucha polipeptydowego. tRNA z aminokwasami, zgodnie z zasadą komplementarności antykodon-kodon, łączy się z mRNA i wchodzi do rybosomu. W centrum peptydowym rybosomu tworzy się wiązanie peptydowe pomiędzy dwoma aminokwasami, a uwolniony tRNA opuszcza rybosom. W tym przypadku mRNA za każdym razem przesuwa się o jedną trójkę, wprowadzając nowy tRNA - aminokwas i usuwając uwolnione tRNA z rybosomu. Cały proces zapewnia energia ATP. Jeden mRNA może łączyć się z kilkoma rybosomami, tworząc polisom, w którym jednocześnie syntetyzowanych jest wiele cząsteczek jednego białka. Synteza kończy się, gdy w mRNA zaczynają się kodony nonsensowne (kody stop). Rybosomy oddzielają się od mRNA i usuwają z nich łańcuchy polipeptydowe. Ponieważ cały proces syntezy odbywa się na ziarnistej siateczce śródplazmatycznej, powstałe łańcuchy polipeptydowe przedostają się do kanalików ER, gdzie uzyskują ostateczną strukturę i przekształcają się w cząsteczki białka.
Wszystkie reakcje syntezy są katalizowane przez specjalne enzymy przy wydatku energii ATP. Szybkość syntezy jest bardzo wysoka i zależy od długości polipeptydu. Na przykład w rybosomie Escherichia coli białko składające się z 300 aminokwasów jest syntetyzowane w ciągu około 15-20 sekund.
Interakcja i struktura mRNA, tRNA, RRNA – trzech głównych kwasów nukleinowych – jest przedmiotem badań cytologii. Pomoże to wyjaśnić rolę transportowego kwasu rybonukleinowego (tRNA) w komórkach. Ta bardzo mała, ale jednocześnie niezaprzeczalnie ważna cząsteczka bierze udział w procesie łączenia białek tworzących organizm.
Jaka jest struktura tRNA? Bardzo interesujące jest spojrzenie na tę substancję „od środka”, aby poznać jej biochemię i rolę biologiczną. A także, w jaki sposób struktura tRNA i jego rola w syntezie białek są ze sobą powiązane?
Co to jest tRNA i jaka jest jego struktura?
Transport kwasu rybonukleinowego bierze udział w budowie nowych białek. Prawie 10% wszystkich kwasów rybonukleinowych to kwasy transportowe. Aby było jasne, z jakich pierwiastków chemicznych zbudowana jest cząsteczka, opiszemy strukturę struktury drugorzędowej tRNA. Struktura wtórna uwzględnia wszystkie podstawowe wiązania chemiczne między pierwiastkami.
Jest to makrocząsteczka składająca się z łańcucha polinukleotydowego. Zawarte w nim zasady azotowe są połączone wiązaniami wodorowymi. Podobnie jak DNA, RNA składa się z 4 zasad azotowych: adeniny, cytozyny, guaniny i uracylu. W tych związkach adenina jest zawsze kojarzona z uracylem, a guanina, jak zwykle, z cytozyną.
Dlaczego nukleotyd ma przedrostek rybo-? Po prostu wszystkie liniowe polimery, które mają rybozę zamiast pentozy u podstawy nukleotydu, nazywane są rybonukleikami. A transferowy RNA jest jednym z 3 typów właśnie takiego polimeru rybonukleinowego.
Struktura tRNA: biochemia
Przyjrzyjmy się najgłębszym warstwom struktury cząsteczki. Te nukleotydy mają 3 składniki:
- Sacharoza, wszystkie typy RNA obejmują rybozę.
- Kwas fosforowy.
- Zasady azotowe. Są to puryny i pirymidyny.

Zasady azotowe są połączone ze sobą silnymi wiązaniami. Zwyczajowo dzieli się zasady na purynę i pirymidynę.
Puryny to adenina i guanina. Adenina odpowiada nukleotydowi adenylowemu składającemu się z 2 połączonych ze sobą pierścieni. A guanina odpowiada temu samemu „jednopierścieniowemu” nukleotydowi guaniny.
Piramidyny to cytozyna i uracyl. Pirymidyny mają strukturę pojedynczego pierścienia. W RNA nie ma tyminy, ponieważ jest ona zastąpiona pierwiastkiem takim jak uracyl. Należy to zrozumieć przed zwróceniem uwagi na inne cechy strukturalne tRNA.
Rodzaje RNA
Jak widać, struktury tRNA nie można opisać w skrócie. Aby zrozumieć cel cząsteczki i jej prawdziwą strukturę, należy zagłębić się w biochemię. Jakie inne nukleotydy rybosomalne są znane? Istnieją także macierzowe lub informacyjne i rybosomalne kwasy nukleinowe. W skrócie mRNA i RRNA. Wszystkie 3 cząsteczki ściśle ze sobą współpracują w komórce, aby zapewnić organizmowi otrzymanie odpowiednio skonstruowanych globul białkowych.

Nie sposób wyobrazić sobie pracy jednego polimeru bez pomocy 2 innych. Cechy strukturalne tRNA stają się wyraźniejsze, gdy rozważymy je w połączeniu z funkcjami bezpośrednio związanymi z pracą rybosomów.
Struktura mRNA, tRNA i RRNA jest pod wieloma względami podobna. Wszystkie mają rybozę u podstawy. Jednak ich struktura i funkcje są różne.
Odkrycie kwasów nukleinowych
W 1868 roku Szwajcar Johann Miescher odkrył makrocząsteczki w jądrze komórkowym, zwane później nukleinami. Nazwa „nukleina” pochodzi od słowa (jądro) – jądro. Chociaż nieco później odkryto, że u istot jednokomórkowych, które nie mają jądra, substancje te są również obecne. W połowie XX wieku otrzymał Nagrodę Nobla za odkrycie syntezy kwasów nukleinowych.
Funkcje tRNA w syntezie białek
Sama nazwa – transfer RNA – mówi o głównej funkcji cząsteczki. Ten kwas nukleinowy „niesie” ze sobą niezbędny aminokwas wymagany przez rybosomalny RNA do wytworzenia określonego białka.
Cząsteczka tRNA ma kilka funkcji. Pierwsza to rozpoznawanie kodonów mRNA, druga funkcja to dostarczanie elementów budulcowych – aminokwasów do syntezy białek. Niektórzy inni eksperci podkreślają funkcję akceptora. Oznacza to dodanie aminokwasów zgodnie z zasadą kowalencyjną. Enzym taki jak syntaza aminocyl-TRNA pomaga „przyłączyć” ten aminokwas.
Jak struktura tRNA jest powiązana z jego funkcjami? Ten specjalny kwas rybonukleinowy jest zaprojektowany w taki sposób, że po jednej stronie znajdują się zasady azotowe, które są zawsze połączone parami. Są to znane nam elementy - A, U, C, G. Dokładnie 3 „litery” lub zasady azotowe tworzą antykodon - odwrotny zestaw elementów, który oddziałuje z kodonem zgodnie z zasadą komplementarności.
Ta ważna cecha struktury tRNA gwarantuje, że podczas dekodowania matrycowego kwasu nukleinowego nie będzie błędów. Przecież od dokładnej sekwencji aminokwasów zależy, czy aktualnie potrzebne organizmowi białko zostanie prawidłowo zsyntetyzowane.
Cechy konstrukcyjne
Jakie są cechy strukturalne tRNA i jego rola biologiczna? To bardzo stara konstrukcja. Jego wymiary mieszczą się w przedziale od 73 do 93 nukleotydów. Masa cząsteczkowa substancji wynosi 25 000–30 000.
Strukturę drugorzędowej struktury tRNA można analizować badając 5 głównych elementów cząsteczki. Zatem ten kwas nukleinowy składa się z następujących elementów:
- pętla do kontaktu z enzymem;
- pętla kontaktu z rybosomem;
- pętla antykodonowa;
- trzon akceptorowy;
- samego antykodonu.
Izolują także małą pętlę zmienną w strukturze wtórnej. Jedno ramię wszystkich typów tRNA jest takie samo – rdzeń złożony z dwóch reszt cytozyny i jednej reszty adenozyny. W tym momencie następuje połączenie z 1 z 20 dostępnych aminokwasów. Każdy aminokwas ma swój własny enzym, swój własny aminoacylo-tRNA.

Cała informacja szyfrująca strukturę wszystkich kwasów nukleinowych zawarta jest w samym DNA. Struktura TRNA jest prawie identyczna u wszystkich żywych istot na planecie. Oglądany w 2D będzie wyglądał jak liść.
Jeśli jednak spojrzeć trójwymiarowo, cząsteczka przypomina strukturę geometryczną w kształcie litery L. Uważa się to za trzeciorzędową strukturę tRNA. Ale dla ułatwienia nauki zwyczajowo „rozwija się” ją wizualnie. Struktura trzeciorzędowa powstaje w wyniku oddziaływania elementów struktury wtórnej, czyli tych części, które się wzajemnie uzupełniają.
Ramiona lub pierścienie TRNA odgrywają ważną rolę. Na przykład jedno ramię jest wymagane do wiązania chemicznego z określonym enzymem.
Charakterystyczną cechą nukleotydu jest obecność ogromnej liczby nukleozydów. Istnieje ponad 60 rodzajów tych mniejszych nukleozydów.
Struktura TRNA i kodowanie aminokwasów
Wiemy, że antykodon TRNA ma długość 3 cząsteczek. Każdy antykodon odpowiada konkretnemu, „osobistemu” aminokwasowi. Aminokwas ten jest połączony z cząsteczką tRNA za pomocą specjalnego enzymu. Gdy tylko 2 aminokwasy się połączą, wiązania z tRNA zostają zerwane. Wszystkie związki chemiczne i enzymy są potrzebne przed wymaganym czasem. W ten sposób struktura i funkcje tRNA są ze sobą powiązane.
W sumie w komórce występuje 61 rodzajów takich cząsteczek. Różnic matematycznych może być 64. Brakuje jednak 3 typów tRNA, ponieważ dokładnie taka sama liczba kodonów stop w mRNA nie ma antykodonów.
Oddziaływanie mRNA i tRNA
Rozważmy interakcję substancji z mRNA i RRNA, a także cechy strukturalne tRNA. Struktura i przeznaczenie makrocząsteczki są ze sobą powiązane.
Struktura mRNA kopiuje informację z oddzielnej części DNA. Sam DNA jest zbyt dużym związkiem cząsteczek i nigdy nie opuszcza jądra. Dlatego potrzebujemy pośredniego RNA – RNA informacyjnego.

W oparciu o sekwencję cząsteczek skopiowaną przez mRNA rybosom buduje białko. Rybosom jest odrębną strukturą polinukleotydową, której budowa wymaga wyjaśnienia.
Rybosomalny tRNA: interakcja
Rybosomalny RNA to ogromna organella. Jego masa cząsteczkowa wynosi 1 000 000 - 1 500 000. Prawie 80% całkowitej ilości RNA stanowią nukleotydy rybosomalne.

Wydaje się, że wychwytuje łańcuch mRNA i czeka na antykodony, które przyniosą ze sobą cząsteczki tRNA. Rybosomalny RNA składa się z 2 podjednostek: małej i dużej.
Rybosom nazywany jest „fabryką”, ponieważ w tej organelli odbywa się cała synteza substancji niezbędnych do życia codziennego. Jest to również bardzo stara struktura komórkowa.
Jak zachodzi synteza białek w rybosomie?
Struktura tRNA i jego rola w syntezie białek są ze sobą powiązane. Antykodon znajdujący się po jednej stronie kwasu rybonukleinowego ma odpowiedni kształt do swojej głównej funkcji - dostarczania aminokwasów do rybosomu, gdzie następuje stopniowa budowa białka. Zasadniczo tRNA działa jako pośrednik. Jego zadaniem jest jedynie dostarczenie niezbędnego aminokwasu.
Kiedy informacja jest odczytywana z jednej części mRNA, rybosom przemieszcza się dalej wzdłuż łańcucha. Matryca jest potrzebna jedynie do przekazania zakodowanej informacji o konfiguracji i funkcji pojedynczego białka. Następnie inny tRNA z zasadami azotowymi zbliża się do rybosomu. Dekoduje także kolejną część mRNA.
Dekodowanie odbywa się w następujący sposób. Zasady azotowe łączą się zgodnie z zasadą komplementarności w taki sam sposób, jak w samym DNA. W związku z tym TRNA widzi, gdzie musi „zacumować” i do którego „hangaru” wysłać aminokwas.

Następnie w rybosomie tak wybrane aminokwasy łączą się chemicznie, krok po kroku powstaje nowa liniowa makrocząsteczka, która po zakończeniu syntezy zostaje skręcona w globulę (kulkę). Zużyte tRNA i mRNA, spełniając swoją funkcję, są usuwane z „fabryki” białka.
Kiedy pierwsza część kodonu łączy się w parę z antykodonem, określana jest ramka odczytu. Następnie, jeśli z jakiegoś powodu nastąpi przesunięcie ramki, wówczas pewna cecha białka zostanie odrzucona. Rybosom nie może interweniować w tym procesie i rozwiązać problemu. Dopiero po zakończeniu procesu 2 podjednostki RRNA ponownie łączą się. Średnio na każde 104 aminokwasów przypada 1 błąd. Na każde 25 już złożonych białek z pewnością wystąpi co najmniej 1 błąd replikacji.
TRNA jako cząsteczki reliktowe
Ponieważ tRNA mogło istnieć w momencie powstania życia na Ziemi, nazywa się je cząsteczką reliktową. Uważa się, że RNA jest pierwszą strukturą, która istniała przed DNA, a następnie ewoluowała. Hipoteza świata RNA została sformułowana w 1986 roku przez laureata Waltera Gilberta. Jednak nadal trudno to udowodnić. Teorię potwierdzają oczywiste fakty - cząsteczki tRNA są w stanie przechowywać bloki informacji i w jakiś sposób tę informację realizować, czyli wykonywać pracę.
Przeciwnicy tej teorii argumentują jednak, że krótki okres życia substancji nie gwarantuje, że tRNA będzie dobrym nośnikiem jakiejkolwiek informacji biologicznej. Te nukleotydy szybko się rozpadają. Żywotność tRNA w komórkach ludzkich waha się od kilku minut do kilku godzin. Niektóre typy mogą trwać do jednego dnia. A jeśli mówimy o tych samych nukleotydach u bakterii, to ramy czasowe są znacznie krótsze – do kilku godzin. Ponadto struktura i funkcje tRNA są zbyt złożone, aby cząsteczka mogła stać się podstawowym elementem biosfery Ziemi.
Wszystkie tRNA mają wspólne cechy zarówno pod względem struktury pierwszorzędowej, jak i sposobu, w jaki łańcuch polinukleotydowy jest składany w strukturę drugorzędową w wyniku interakcji pomiędzy zasadami reszt nukleotydowych.
Podstawowa struktura tRNA
tRNA są stosunkowo małymi cząsteczkami, długość ich łańcuchów waha się od 74 do 95 reszt nukleotydowych. Wszystkie tRNA mają ten sam koniec 3", zbudowany z dwóch reszt cytozyny i jednej reszty adenozyny (koniec CCA). To 3"-końcowa adenozyna wiąże się z resztą aminokwasu podczas tworzenia aminoacylo-tRNA. Koniec CCA jest przyłączony do wielu tRNA za pomocą specjalnego enzymu. Triplet nukleotydowy komplementarny do kodonu aminokwasu (antykodon) znajduje się mniej więcej pośrodku łańcucha tRNA. W pewnych pozycjach sekwencji prawie wszystkie typy tRNA zawierają te same (konserwatywne) reszty nukleotydowe. Niektóre pozycje mogą zawierać tylko zasady purynowe lub tylko pirymidynowe (nazywa się je resztami semikonserwatywnymi).
Wszystkie cząsteczki tRNA charakteryzują się obecnością dużej liczby (do 25% wszystkich reszt) różnych modyfikowanych nukleozydów, często nazywanych drobnymi. Powstają w różnych miejscach cząsteczek, w wielu przypadkach dobrze zdefiniowanych, w wyniku modyfikacji zwykłych reszt nukleozydowych przez specjalne enzymy.
Struktura drugorzędowa tRNA
złożenie łańcucha w strukturę wtórną następuje na skutek wzajemnego uzupełniania się odcinków łańcucha. Trzy fragmenty łańcucha uzupełniają się, gdy są złożone na sobie, tworząc struktury przypominające spinkę do włosów. Ponadto koniec 5" jest komplementarny w stosunku do obszaru blisko końca 3" łańcucha, z ich ułożeniem antyrównoległym; tworzą tzw. rdzeń akceptorowy. Rezultatem jest struktura charakteryzująca się obecnością czterech łodyg i trzech pętli, zwana „liścią koniczyny”. Trzon i pętla tworzą gałąź. Na dole znajduje się gałąź antykodonu, która zawiera triplet antykodonu jako część swojej pętli. Po lewej i prawej stronie znajdują się gałęzie D i T, nazwane odpowiednio ze względu na obecność w swoich pętlach niezwykłych konserwatywnych nukleozydów, dihydrourydyny (D) i tymidyny (T). Sekwencje nukleotydowe wszystkich badanych tRNA można złożyć w podobne struktury. Oprócz trzech pętli koniczyny, tRNA ma również dodatkową lub zmienną pętlę (pętlę V). Jego rozmiary różnią się znacznie w przypadku różnych tRNA i wahają się od 4 do 21 nukleotydów, a według najnowszych danych do 24 nukleotydów.
Przestrzenna (trzeciorzędowa) struktura tRNA
W wyniku interakcji elementów struktury wtórnej powstaje struktura trzeciorzędowa, zwana formą L ze względu na podobieństwo do łacińskiej litery L (ryc. 2 i 3). Dzięki ułożeniu zasad łodyga akceptorowa i łodyga T koniczyny tworzą jedną ciągłą podwójną helisę, a pozostałe dwie łodygi, antykodon i D, tworzą kolejną ciągłą podwójną helisę. W tym przypadku pętle D i T są zbliżane do siebie i łączone ze sobą poprzez utworzenie dodatkowych, często nietypowych par zasad. W tworzeniu tych par z reguły biorą udział reszty konserwatywne lub półkonserwatywne. Podobne interakcje trzeciorzędowe spajają niektóre inne części struktury L
Rybosomalny RNA
Rybosomalne kwasy rybonukleinowe (rRNA) to kilka cząsteczek RNA, które stanowią podstawę rybosomu. Główną funkcją rRNA jest przeprowadzanie procesu translacji - odczytywanie informacji z mRNA za pomocą cząsteczek adaptorowych tRNA i katalizowanie tworzenia wiązań peptydowych pomiędzy aminokwasami przyłączonymi do tRNA. Rybosomalny RNA stanowi około 80% całkowitego RNA komórki. Jest kodowany przez geny znajdujące się w DNA kilku chromosomów zlokalizowanych w obszarze jąderka zwanym organizatorem jąderkowym.
Sekwencja zasad w rRNA jest podobna we wszystkich organizmach, od bakterii po zwierzęta. rRNA znajduje się w cytoplazmie, gdzie wiąże się z cząsteczkami białek, tworząc razem organelle komórkowe zwane rybosomami. Synteza białek zachodzi na rybosomach. Tutaj „kod” zawarty w mRNA ulega translacji na sekwencję aminokwasów łańcucha polipeptydowego.
Przenieść RNA
Transferowy RNA, tRNA, to kwas rybonukleinowy, którego funkcją jest transport aminokwasów do miejsca syntezy białek. tRNA biorą także bezpośredni udział w wydłużaniu łańcucha polipeptydowego, łącząc się – będąc w kompleksie z aminokwasem – z kodonem mRNA i zapewniając złożoną konformację niezbędną do utworzenia nowego wiązania peptydowego.
Każdy aminokwas ma swój własny tRNA.
tRNA jest jednoniciowym RNA, ale w swojej funkcjonalnej formie ma konformację liścia koniczyny. Składa się z czterech głównych części, które spełniają różne funkcje. „Rdzeń” akceptorowy jest utworzony przez dwie komplementarne, połączone końcowe części tRNA. Składa się z siedmiu par zasad. 3-calowy koniec tego rdzenia jest nieco dłuższy i tworzy region jednoniciowy zakończony sekwencją CCA z wolną grupą OH. Do tego końca jest przyłączony transportowany aminokwas. Pozostałe trzy gałęzie to komplementarne sparowane sekwencje nukleotydowe, które kończą się w niesparowanych regionach tworzących pętle.Środkowa z tych gałęzi – antykodon – składa się z pięciu par nukleotydów i zawiera antykodon w środku swojej pętli.Antykodon to trzy nukleotydy komplementarne do kodonu mRNA, który koduje transportowany aminokwas przez ten tRNA do miejsca syntezy peptydu.
Pomiędzy gałęziami akceptorowymi i antykodonowymi znajdują się dwie gałęzie boczne. W swoich pętlach zawierają zmodyfikowane zasady -dihydrourydynę (pętla D) i triplet T?C, gdzie? - pseudourain (pętla T?C). Pomiędzy gałęziami aitikodonu i TΔC znajduje się dodatkowa pętla zawierająca od 3-5 do 13-21 nukleotydów.
Aminokwas jest kowalencyjnie przyłączony do 3" końca cząsteczki za pomocą enzymu syntetazy aminoacylo-tRNA, specyficznego dla każdego typu tRNA.
tRNA służy jako cząsteczka pośrednia pomiędzy kodonem tripletowym w mRNA a sekwencją aminokwasów łańcucha polipeptydowego. tRNA stanowi około 15% całego komórkowego RNA; te RNA mają najkrótszy łańcuch polinukleotydowy – zawiera średnio 80 nukleotydów. Każda pojedyncza komórka zawiera ponad 20 różnych cząsteczek tRNA. Wszystkie cząsteczki tRNA mają podobną podstawową strukturę. Na końcu 5' cząsteczki tRNA zawsze znajduje się guanina, a na końcu 3' sekwencja zasad CCA.
Sekwencja nukleotydów w pozostałej części cząsteczki jest zmienna i może zawierać „niezwykłe” zasady, takie jak inozyna i pseudouracyl.
Sekwencja zasad w triplecie antykodonu ściśle odpowiada aminokwasowi zawartemu w tej cząsteczce tRNA.
Ryż. 3.
Każdy aminokwas jest przyłączony do jednego ze swoich specyficznych tRNA przy udziale enzymu syntazy aminoacylo-tRNA. Powoduje to utworzenie kompleksu animokwas-tRNA, znanego jako animoacylo-tRNA, w którym energia wiązania między końcowym nukleotydem A w triplecie CCA a aminokwasem jest wystarczająca, aby umożliwić późniejsze wiązanie z sąsiednim aminokwasem. W ten sposób syntetyzowany jest łańcuch polipeptydowy.
Jedną z cech tRNA jest obecność w nim nietypowych zasad, które powstają w wyniku modyfikacji chemicznej po włączeniu normalnej zasady do łańcucha polinukleotydowego. Te zmienione zasady determinują dużą różnorodność strukturalną tRNA w ogólnym planie ich struktury. Najbardziej interesujące są modyfikacje zasad tworzących antykodon, które wpływają na specyficzność jego interakcji z kodonem. Na przykład atypowa zasada inozyna, czasami znajdująca się na 1. pozycji antykodonu tRNA, jest zdolna do komplementarnego łączenia się z trzema różnymi trzecimi zasadami kodonu mRNA - U, C i A. Ponieważ jedną z cech kodu genetycznego jest ze względu na degenerację wiele aminokwasów jest szyfrowanych przez kilka kodonów, które zazwyczaj różnią się trzecią zasadą. Ze względu na niespecyficzne wiązanie zmodyfikowanej zasady antykodonu, jeden tRNA rozpoznaje kilka synonimicznych kodonów.