RNA sekundaarne struktuur. Ribonukleiinhapete struktuur ja funktsioonid
RNA molekul on samuti polümeer, mille monomeerideks on ribonukleotiidid, RNA on üheahelaline molekul. See on üles ehitatud samamoodi nagu üks DNA ahelatest. RNA nukleotiidid on sarnased DNA nukleotiididega, kuigi nad ei ole nendega identsed. Neid on samuti neli ja need koosnevad lämmastikaluse, pentoosi ja fosforhappe jääkidest. Kolm lämmastikualust on täpselt samad, mis DNA-s: A, G ja C. Siiski selle asemel T RNA-s sisalduv DNA sisaldab sarnase struktuuriga pürimidiini alust uratsiili ( Kell). Peamine erinevus DNA ja RNA vahel on süsivesikute olemus: DNA nukleotiidides on monosahhariid desoksüriboos ja RNA-s riboos. Nukleotiidide vaheline ühendus toimub nagu DNA-s suhkru ja fosforhappe jäägi kaudu. Erinevalt DNA-st, mille sisaldus on teatud organismide rakkudes konstantne, on RNA sisaldus neis kõikuv. See on märgatavalt kõrgem seal, kus toimub intensiivne süntees.
Seoses täidetavate funktsioonidega eristatakse mitut tüüpi RNA-d.
RNA ülekandmine (tRNA). tRNA molekulid on kõige lühemad: koosnevad vaid 80-100 nukleotiidist. Selliste osakeste molekulmass on 25-30 tuhat Transpordi-RNA-d sisalduvad peamiselt raku tsütoplasmas. Nende ülesanne on viia aminohapped ribosoomidesse, valgusünteesi kohta. Rakkude RNA kogusisaldusest moodustab tRNA umbes 10%.
Ribosomaalne RNA (rRNA). Need on suured molekulid: need sisaldavad vastavalt 3-5 tuhat nukleotiidi, nende molekulmass ulatub 1-1,5 miljonini.Ribosomaalsed RNA-d moodustavad ribosoomi olulise osa. Kogu RNA sisaldusest rakus moodustab rRNA umbes 90%.
Messenger RNA (mRNA) või sõnumitooja RNA (mRNA), mida leidub tuumas ja tsütoplasmas. Selle ülesanne on edastada teavet valgu struktuuri kohta DNA-st ribosoomide valgusünteesi kohta. mRNA osakaal moodustab ligikaudu 0,5-1% kogu raku RNA sisaldusest. mRNA suurus on väga erinev - 100 kuni 10 000 nukleotiidi.
Kõik RNA tüübid sünteesitakse DNA-l, mis toimib omamoodi mallina.
DNA on päriliku teabe kandja.
Iga valku esindab üks või mitu polüpeptiidahelat. DNA osa, mis kannab teavet ühe polüpeptiidahela kohta, nimetatakse genoom. DNA molekulide kogum rakus toimib geneetilise informatsiooni kandjana. Geneetiline teave edastatakse emarakkudelt tütarrakkudele ja vanematelt lastele. Geen on geneetika ühik, või pärilikku teavet.
DNA on rakus geneetilise informatsiooni kandja - ei osale otseselt valkude sünteesis. Eukarüootsetes rakkudes sisalduvad DNA molekulid tuuma kromosoomides ja eraldatakse tuumamembraaniga tsütoplasmast, kus sünteesitakse valke. Ribosoomidesse - valkude kogunemiskohtadesse - saadetakse tuumast infokandja, mis on võimeline läbima tuumaümbrise poore. Messenger RNA (mRNA) on selline vahendaja. Komplementaarsuse põhimõtte kohaselt sünteesitakse see DNA-l ensüümi RNA- osalusel. polümeraas.

Messenger RNA on üheahelaline molekul ja transkriptsioon pärineb kaheahelalise DNA molekuli ühest ahelast. See ei ole kogu DNA molekuli koopia, vaid ainult osa sellest - üks geen eukarüootides või külgnevate geenide rühm, mis kannab teavet valkude struktuuri kohta, mis on vajalikud ühe funktsiooni täitmiseks prokarüootides. Seda geenide rühma nimetatakse operon. Iga operoni alguses on omamoodi RNA polümeraasi maandumiskoht, mida nimetatakse promootor.see on spetsiifiline DNA nukleotiidide järjestus, mille ensüüm "ära tunneb" keemilise afiinsuse tõttu. Ainult promootori külge kinnitumisel on RNA polümeraas võimeline käivitama RNA sünteesi. Olles jõudnud operoni lõppu, kohtab ensüüm signaali (teatud nukleotiidide järjestuse kujul), mis näitab lugemise lõppu. Valmis mRNA eemaldub DNA-st ja läheb valgusünteesi kohta.
Transkriptsiooniprotsessis on neli etappi: 1) RNA sidumine- polümeraas koos promootoriga; 2) algatus- sünteesi algus. See seisneb esimese fosfodiestersideme moodustamises ATP või GTP ja sünteesitud RNA molekuli teise nukleotiidi vahel; 3) pikenemine– RNA ahela kasv; need. nukleotiidide järjestikune lisamine üksteisele selles järjekorras, milles nende komplementaarsed nukleotiidid on transkribeeritud DNA ahelas. Pikenduskiirus on 50 nukleotiidi sekundis; 4) lõpetamine- RNA sünteesi lõpetamine.
Pärast tuumaümbrise pooride läbimist saadetakse mRNA ribosoomidesse, kus dešifreeritakse geneetiline informatsioon – see tõlgitakse nukleotiidide "keelest" aminohapete "keelde". Ribosoomides esinev polüpeptiidahelate süntees mRNA matriitsi järgi on nn. saade(lat. tõlge - tõlge).
Aminohapped, millest sünteesitakse valke, viiakse ribosoomidesse spetsiaalsete RNA-de abil, mida nimetatakse transport-RNA-deks (tRNA). Erinevaid tRNA-sid on rakus sama palju kui aminohappeid kodeerivaid koodoneid. Iga tRNA "lehe" ülaosas on kolmest nukleotiidist koosnev järjestus, mis on komplementaarsed mRNA koodoni nukleotiididega. Nad kutsuvad teda antikoodon. Spetsiaalne ensüüm – kodaas – tunneb ära tRNA ja kinnitab "lehelehele" aminohappe – ainult selle, mida kodeerib antikoodoniga komplementaarne triplet. Ühe ATP molekuli energia kulutatakse kovalentse sideme moodustamiseks tRNA ja selle "oma" aminohappe vahel.
 Selleks, et aminohape sisalduks polüpeptiidahelas, peab see tRNA-st lahku lööma. See saab võimalikuks, kui tRNA siseneb ribosoomi ja antikoodon tunneb mRNA-s ära oma koodoni. Ribosoomil on kaks kohta kahe tRNA molekuli sidumiseks. Üks neist valdkondadest, nn aktsepteerija tRNA siseneb koos aminohappega ja kinnitub selle koodoniga (I). Kas see aminohape kinnitub enda külge (aktsepteerib) kasvavat valguahelat (II)? Nende vahel moodustub peptiidside. tRNA, mis on nüüd ühendatud koos mRNA koodoniga doonor ribosoomi osa. Vabanenud aktseptorsaiti tuleb uus tRNA, mis on seotud aminohappega, mille krüpteerib järgmine koodon (III). Doonorkohalt kantakse eraldunud polüpeptiidahel uuesti üle siia ja pikendatakse veel ühe lüli võrra. Kasvavas ahelas olevad aminohapped on ühendatud järjestuses, milles mRNA-s asuvad neid kodeerivad koodonid.
Selleks, et aminohape sisalduks polüpeptiidahelas, peab see tRNA-st lahku lööma. See saab võimalikuks, kui tRNA siseneb ribosoomi ja antikoodon tunneb mRNA-s ära oma koodoni. Ribosoomil on kaks kohta kahe tRNA molekuli sidumiseks. Üks neist valdkondadest, nn aktsepteerija tRNA siseneb koos aminohappega ja kinnitub selle koodoniga (I). Kas see aminohape kinnitub enda külge (aktsepteerib) kasvavat valguahelat (II)? Nende vahel moodustub peptiidside. tRNA, mis on nüüd ühendatud koos mRNA koodoniga doonor ribosoomi osa. Vabanenud aktseptorsaiti tuleb uus tRNA, mis on seotud aminohappega, mille krüpteerib järgmine koodon (III). Doonorkohalt kantakse eraldunud polüpeptiidahel uuesti üle siia ja pikendatakse veel ühe lüli võrra. Kasvavas ahelas olevad aminohapped on ühendatud järjestuses, milles mRNA-s asuvad neid kodeerivad koodonid.
Kui ribosoomilt leitakse üks kolmest kolmikust ( UAA, UAG, UGA), mis on geenidevahelised "kirjavahemärgid", ei saa ükski tRNA aktseptorsaidis asuda. Fakt on see, et puuduvad antikoodonid, mis oleksid komplementaarsed "kirjavahemärkide" nukleotiidjärjestustega. Eraldunud ahelal pole aktseptorkohas midagi külge kinnituda ja see lahkub ribosoomist. Valkude süntees on lõppenud.
Prokarüootides algab valgusüntees koodonist AUG, mis asub iga geeni koopias esimesel kohal, on ribosoomis sellisel positsioonil, et spetsiaalse tRNA antikoodon interakteerub sellega, mis on seotud formüülmentioniin. See aminohappe metioniini modifitseeritud vorm siseneb koheselt doonorikohta ja täidab fraasis suure algustähe rolli - mis tahes polüpeptiidahela süntees algab sellega bakterirakus. Kui kolmik AUG ei ole esiteks, vaid geeni koopia sees kodeerib see aminohapet metioniini. Pärast polüpeptiidahela sünteesi lõppu eraldatakse formüülmetioniin sellest ja seda valmisvalgus ei leidu.
 Valkude tootmise suurendamiseks läbib mRNA sageli korraga mitte ühte, vaid mitut ribosoomi. Millist struktuuri, mida ühendab üks mRNA molekul, nimetatakse polüsoom. Igal ribosoomil sünteesitakse sellel helmelaadsel konveieril identsed valgud.
Valkude tootmise suurendamiseks läbib mRNA sageli korraga mitte ühte, vaid mitut ribosoomi. Millist struktuuri, mida ühendab üks mRNA molekul, nimetatakse polüsoom. Igal ribosoomil sünteesitakse sellel helmelaadsel konveieril identsed valgud.
Aminohapped tarnitakse ribosoomidesse pidevalt tRNA kaudu. Pärast aminohappe annetamist lahkub tRNA ribosoomist ja ühendatakse kodaasi abil. Kõigi valkude tootmiseks vajalike "tehase teenuste" kõrge sidusus võimaldab mõne sekundi jooksul sünteesida sadadest aminohapetest koosnevaid polüpeptiidahelaid.
Geneetilise koodi omadused. Rakus toimuva transkriptsiooniprotsessi kaudu kantakse teave DNA-st üle valku.
DNA → mRNA → valk
DNA-s ja mRNA-s sisalduv geneetiline informatsioon sisaldub molekulide nukleotiidide järjestuses.
 Kuidas toimub informatsiooni tõlkimine nukleotiidide "keelest" aminohapete "keelde"? See tõlge viiakse läbi geneetilise koodi abil. kood või šifr, on sümbolite süsteem ühe teabevormi teiseks tõlkimiseks. Geneetiline kood on süsteem teabe salvestamiseks valkude aminohapete järjestuse kohta, kasutades mRNA nukleotiidide järjestust.
Kuidas toimub informatsiooni tõlkimine nukleotiidide "keelest" aminohapete "keelde"? See tõlge viiakse läbi geneetilise koodi abil. kood või šifr, on sümbolite süsteem ühe teabevormi teiseks tõlkimiseks. Geneetiline kood on süsteem teabe salvestamiseks valkude aminohapete järjestuse kohta, kasutades mRNA nukleotiidide järjestust.
Millised on geneetilise koodi omadused?
kolmiku kood. RNA sisaldab nelja nukleotiidi: A, G, C, W. Kui prooviksime määrata ühte aminohapet ühe nukleotiidiga, jääks 16 aminohapet 20-st krüptimata. Kahetäheline kood krüpteerib 16 aminohapet. Loodus on loonud kolmetähelise ehk kolmiku koodi. See tähendab et iga 20 aminohappest on kodeeritud kolmest nukleotiidist koosneva järjestusega, mida nimetatakse tripletiks või koodoniks.
Kood on degenereerunud. See tähendab et iga aminohapet kodeerib rohkem kui üks koodon. Erandid: meteoniin ja trüptofaan, millest igaüks on kodeeritud ühe tripleti poolt.
Kood on üheselt mõistetav. Iga koodon kodeerib ainult ühte aminohapet.
Geenide vahel on "kirjavahemärgid". Trükitekstis on iga fraasi lõpus punkt. Mitmed seotud fraasid moodustavad lõigu. Geneetilise informatsiooni keeles on selline lõik operon ja seda komplementaarne mRNA. Iga geen prokarüootses operonis või üksikus eukarüootses geenis kodeerib ühte polüpeptiidahelat – fraasi. Kuna mõnel juhul luuakse mRNA matriitsile järjestikku mitu erinevat polüpeptiidahelat, tuleb need üksteisest eraldada. Selleks on geneetilisel aastal kolm spetsiaalset kolmikut - UAA, UAG, UGA, millest igaüks näitab ühe polüpeptiidahela sünteesi katkemist. Seega täidavad need kolmikud kirjavahemärkide funktsiooni. Need on iga geeni lõpus.
Geeni sees pole "vahemärke".
Kood on universaalne. Geneetiline kood on kõigi Maal elavate olendite jaoks sama. Bakterites ja seentes, nisus ja puuvillas, kalades ja ussides, konnades ja inimestes kodeerivad samad kolmikud samu aminohappeid.

DNA replikatsiooni põhimõtted. Protsess tagab geneetilise materjali järjepidevuse rakkude ja organismide põlvkondades replikatsioon – DNA molekulide dubleerimine. Seda keerukat protsessi viib läbi mitme ensüümi ja valgu kompleks, millel puudub katalüütiline aktiivsus ja mis on vajalikud polünukleotiidahelatele soovitud konformatsiooni saamiseks. Replikatsiooni tulemusena moodustuvad kaks identset DNA kaksikheeliksit. Need niinimetatud tütarmolekulid ei erine üksteisest ega algsest DNA algmolekulist. Replikatsioon toimub rakus enne jagunemist, seega saab iga tütarrakk täpselt samad DNA molekulid, mis emarakul. Replikatsiooniprotsess põhineb mitmel põhimõttel:
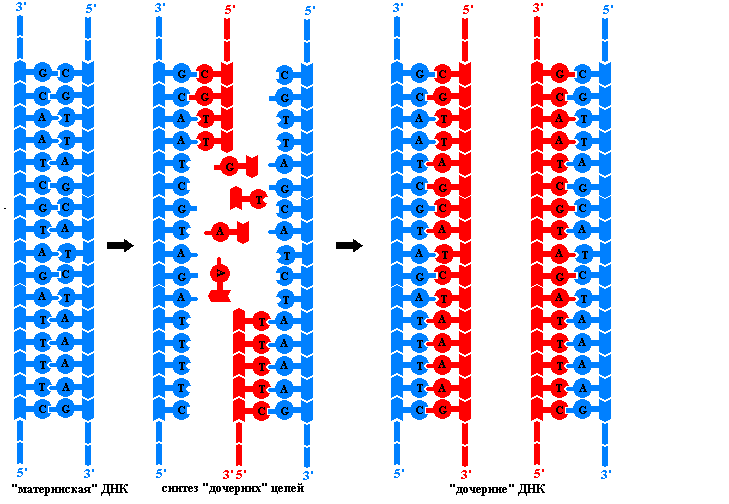
Ainult sel juhul on DNA polümeraasid võimelised liikuma mööda lähteahelaid ja kasutama neid tütarahelate veatu sünteesi mallidena. Kuid paljudest miljonitest aluspaaridest koosnevate spiraalide täielik lahtikerimine on seotud nii märkimisväärse arvu pöörete ja selliste energiakuludega, mis rakutingimustes on võimatud. Seetõttu algab replikatsioon eukarüootides DNA molekuli mõnes kohas samaaegselt. Nimetatakse piirkonda kahe punkti vahel, kust algab tütarahelate süntees replikon. Ta on replikatsiooni ühik.

Iga eukarüootse raku DNA molekul sisaldab palju replikone. Igas replikonis võib näha replikatsioonikahvlit – seda DNA molekuli osa, mis on spetsiaalsete ensüümide toimel juba lahti harutatud. Iga hargi ahel toimib täiendava tütarahela sünteesi mallina. Replikatsiooni ajal liigub kahvel mööda lähtemolekuli, samas kui uued DNA lõigud on lahti keeratud. Kuna DNA polümeraasid saavad maatriksi ahelaid mööda liikuda ainult ühes suunas ja ahelad on orienteeritud antiparalleelselt, sünteesitakse igas hargis korraga kaks erinevat ensümaatilist kompleksi. Veelgi enam, igas kahvlis kasvab üks tütarahel (juht) pidevalt ja teine (mahajäänud) ahel sünteesitakse mitme nukleotiidi pikkuste eraldi fragmentidega. Sellised ensüümid said nime need avastanud Jaapani teadlase järgi Okazaki killud on seotud DNA ligaasiga, moodustades pideva ahela. DNA fragmentide tütarahelate moodustumise mehhanismi nimetatakse katkendlikuks.
Praimeri DNA polümeraasi vajadus ei suuda käivitada juhtiva ahela sünteesi ega ka mahajäänud ahela Okazaki fragmentide sünteesi. See saab üles ehitada ainult juba olemasoleva polünukleotiidahela, kinnitades järjestikku desoksüribonukleotiide oma 3'-OH-otsa. Kust pärineb kasvava DNA ahela esialgne 5' ots? Seda sünteesib DNA matriitsil spetsiaalne RNA polümeraas, mida nimetatakse primaas(Inglise Primer - seeme). Ribonukleotiidpraimeri suurus on väike (alla 20 nukleotiidi) võrreldes DNA poimeraasi poolt moodustatud DNA ahela suurusega. Täitis oma Funktsioonid RNA praimer eemaldatakse spetsiaalse ensüümi abil ning selle käigus tekkinud tühimiku parandab DNA polümeraas, mis kasutab praimerina naabruses asuva Okazaki fragmendi 3'-OH otsa.
Lineaarsete DNA molekulide otste alareplikatsiooni probleem. Äärmuslike RNA praimerite eemaldamine, mis on komplementaarne lineaarse DNA algmolekuli mõlema ahela 3'-otsadega, viib selleni, et alamahelad on lühemad kui 10-20 nukleotiidi. See on lineaarsete molekulide otste alareplikatsiooni probleem.
Lineaarsete DNA molekulide 3' otste alareplikatsiooni probleemi lahendavad eukarüootsed rakud spetsiaalse ensüümi abil - telomeraas.
Telomeraas on DNA polümeraas, mis täiendab kromosoomide 3'-terminaalseid DNA molekule lühikeste korduvate järjestustega. Need, paiknedes üksteise järel, moodustavad korrapärase kuni 10 tuhande nukleotiidi pikkuse terminaalse struktuuri. Telomeraas sisaldab lisaks valguosale RNA-d, mis toimib matriitsina DNA pikendamisel kordustega.
DNA molekulide otste pikenemise skeem. Esiteks toimub väljaulatuva DNA otsa komplementaarne seondumine telomeraasi RNA matriitsi saidiga, seejärel kogub telomeraas DNA, kasutades seemnena selle 3'-OH otsa ja matriitsina RNA-d, mis on osa ensüümist. Seda etappi nimetatakse pikenemiseks. Pärast seda toimub translokatsioon, s.o. DNA liikumine, mida pikendatakse ühe kordusega, ensüümi suhtes. Sellele järgneb pikenemine ja teine translokatsioon.
Selle tulemusena moodustuvad kromosoomide spetsiaalsed lõppstruktuurid. Need koosnevad korduvalt korduvatest lühikestest DNA järjestustest ja spetsiifilistest valkudest.
rRNA ja tRNA prekursorite süntees on sarnane ire-mRNA sünteesiga. Ribosomaalse RNA esmane transkript ei sisalda introneid ja spetsiifiliste RNaaside toimel lõhustatakse see, moodustades 28S-, 18S- ja 5,8S-pRNA; 5S-pRNA sünteesitakse RNA polümeraas III osalusel.
rRNA ja tRNA.
Primaarsed tRNA transkriptid muundatakse ka osalise hüdrolüüsi teel küpseteks vormideks.
Valkude biosünteesis osalevad kõik RNA tüübid, kuid nende funktsioonid selles protsessis on erinevad. Valkude primaarstruktuuri määrava maatriksi rolli täidavad messenger RNA-d (mRNA-d) Translatsioonimehhanismide uurimisel on suur tähtsus valkude biosünteesi rakuvabade süsteemide kasutamisel. Kui koehomogenaate inkubeeritakse aminohapete seguga, millest vähemalt üks on märgistatud, saab valgu biosünteesi registreerida märgise lisamisega valkudesse. Sünteesitud valgu primaarstruktuuri määrab süsteemi lisatud mRNA primaarstruktuur. Kui rakuvaba süsteem koosneb globiini mRNA-st (seda saab eraldada retikulotsüütidest), sünteesitakse globiini (globiini a- ja (3-ahelad), kui albumiin sünteesitakse hepatotsüütidest eraldatud albumiini mRNA-st jne.
14. Replikatsiooni väärtus:
a) protsess on oluline molekulaarne mehhanism, mis on igat tüüpi proeukarüootsete rakkude jagunemise aluseks, b) tagab nii ühe- kui ka mitmerakuliste organismide igat tüüpi paljunemise,
c) säilitab raku püsivuse
elundite, kudede ja organismi koostis füsioloogilise regeneratsiooni tulemusena
d) tagab üksikisikute pikaajalise eksistentsi;
e) tagab organismiliikide pikaajalise eksisteerimise;
e) protsess aitab kaasa teabe täpsele kahekordistamisele;
g) replikatsiooniprotsessis on võimalikud vead (mutatsioonid), mis võivad patoloogiliste muutuste tekkega põhjustada valgusünteesi häireid.
DNA molekuli ainulaadset omadust kahekordistuda enne rakkude jagunemist nimetatakse replikatsiooniks.
Natiivse DNA kui päriliku teabe kandja eriomadused:
1) replikatsioon - uute ahelate moodustumine on komplementaarne;
2) enesekorrektsioon – DNA polümeraas lõikab ära ekslikult paljunenud piirkonnad (10-6);
3) heastamine - taastamine;
Nende protsesside rakendamine toimub rakus spetsiaalsete ensüümide osalusel.
Kuidas parandussüsteem töötab Eksperimendid, mis paljastasid parandamise mehhanismid ja selle võime olemasolu, viidi läbi ainuraksete organismide abil. Kuid parandusprotsessid on omased loomade ja inimeste elusrakkudele. Mõned inimesed kannatavad xeroderma pigmentosum'i all. See haigus on põhjustatud rakkude võimetusest kahjustatud DNA-d uuesti sünteesida. Xeroderma on pärilik. Millest on tehtud parandussüsteem? Neli parandusprotsessi toetavat ensüümi on DNA helikaas, -eksonukleaas, -polümeraas ja -ligaas. Esimene neist ühenditest suudab ära tunda kahjustusi desoksüribonukleiinhappe molekuli ahelas. See mitte ainult ei tunne ära, vaid ka lõikab ahela õiges kohas ära, et eemaldada molekuli muutunud segment. Elimineerimine ise toimub DNA eksonukleaasi abil. Järgmisena sünteesitakse aminohapetest uus desoksüribonukleiinhappemolekuli segment, et kahjustatud segment täielikult asendada. Noh, selle kõige keerukama bioloogilise protseduuri viimane akord viiakse läbi ensüümi DNA ligaasi abil. See vastutab sünteesitud saidi kinnitamise eest kahjustatud molekuliga. Pärast seda, kui kõik neli ensüümi on oma töö teinud, uueneb DNA molekul täielikult ja kõik kahjustused on minevik. Nii toimivad elusraku sees olevad mehhanismid harmoonias.
Klassifikatsioon Praegu eristavad teadlased järgmist tüüpi parandussüsteeme. Need aktiveeritakse sõltuvalt erinevatest teguritest. Nende hulka kuuluvad: Taasaktiveerimine. rekombinatsiooni taastumine. Heteroduplekside remont. ekstsisiooni remont. DNA molekulide mittehomoloogsete otste taasühinemine. Kõigil üherakulistel organismidel on vähemalt kolm ensüümisüsteemi. Igaüks neist on võimeline taastamisprotsessi läbi viima. Nende süsteemide hulka kuuluvad: otsene, ekstsisioon- ja replikatiivne. Prokarüootidel on need kolm DNA parandamise tüüpi. Mis puudutab eukarüoote, siis nende käsutuses on täiendavad mehhanismid, mida nimetatakse Miss-Mathe ja Sos-repair. Bioloogia on üksikasjalikult uurinud kõiki seda tüüpi rakkude geneetilise materjali iseparanemist.
15. Geneetiline kood on meetod valkude aminohappejärjestuse kodeerimiseks, kasutades nukleotiidide järjestust, mis on iseloomulik kõigile elusorganismidele. Valgumolekulis olev aminohappejärjestus krüpteeritakse DNA molekulis nukleotiidjärjestusena ja seda nimetatakse geneetiline kood. DNA molekuli piirkonda, mis vastutab ühe valgu sünteesi eest, nimetatakse genoom.
DNA-s kasutatakse nelja nukleotiidi - adeniini (A), guaniini (G), tsütosiini (C), tümiini (T), mida venekeelses kirjanduses tähistatakse tähtedega A, G, C ja T. Need tähed moodustavad geneetilise koodi tähestik. RNA-s kasutatakse samu nukleotiide, välja arvatud tümiin, mis on asendatud sarnase nukleotiidiga - uratsiiliga, mida tähistatakse tähega U (venekeelses kirjanduses U). DNA ja RNA molekulides reastuvad nukleotiidid ahelatesse ja nii saadakse geneetiliste tähtede järjestused.
Looduses kasutatakse valkude ehitamiseks 20 erinevat aminohapet. Iga valk on ahel või mitu aminohapete ahelat rangelt määratletud järjestuses. See järjestus määrab valgu struktuuri ja seega kõik selle bioloogilised omadused. Aminohapete komplekt on universaalne ka peaaegu kõigile elusorganismidele.
Geneetilise informatsiooni rakendamine elusrakkudes (st geeni poolt kodeeritud valgu süntees) toimub kahe maatriksprotsessi abil: transkriptsioon (st mRNA süntees DNA matriitsil) ja geneetilise koodi translatsioon aminohappeks. järjestus (polüpeptiidahela süntees mRNA matriitsil). 20 aminohappe kodeerimiseks piisab kolmest järjestikusest nukleotiidist ja ka stoppsignaalist, mis tähendab valgujärjestuse lõppu. Kolmest nukleotiidist koosnevat komplekti nimetatakse tripletiks. Aminohapetele ja koodonitele vastavad aktsepteeritud lühendid on näidatud joonisel.
Geneetilise koodi omadused
Kolmilisus – oluline koodiühik on kolme nukleotiidi kombinatsioon (triplet ehk koodon).
Järjepidevus - kolmikute vahel ei ole kirjavahemärke, see tähendab, et teavet loetakse pidevalt.
Mittekattuv – sama nukleotiid ei saa olla samaaegselt kahe või enama kolmiku osa. (Ei kehti mõnede kattuvate geenide kohta viirustes, mitokondrites ja bakterites, mis kodeerivad mitut kaadrinihke valku.)
Ühetähenduslikkus – teatud koodon vastab ainult ühele aminohappele. (Omadus ei ole universaalne. Euplotes crassuse UGA koodon kodeerib kahte aminohapet, tsüsteiini ja selenotsüsteiini)
Degeneratsioon (redundantsus) – samale aminohappele võib vastata mitu koodonit.
Universaalsus - geneetiline kood töötab erineva keerukusega organismides ühtemoodi - viirustest inimeseni (sellel põhinevad geenitehnoloogia meetodid) (Sellel omadusel on ka mitmeid erandeid, vt tabelit "Variatsioonid" standardse geneetilise koodi osa" käesolevas artiklis).
16.Biosünteesi tingimused
Valkude biosüntees nõuab DNA molekuli geneetilist teavet; informatiivne RNA - selle teabe kandja tuumast sünteesikohta; ribosoomid - organellid, kus toimub tegelik valgusüntees; aminohapete komplekt tsütoplasmas; aminohappeid kodeerivate RNA-de transportimine ja nende viimine ribosoomidel sünteesikohta; ATP on aine, mis annab energiat kodeerimise ja biosünteesi protsesside jaoks.
Etapid
Transkriptsioon- igat tüüpi RNA biosünteesi protsess DNA maatriksil, mis toimub tuumas.
Teatud osa DNA molekulist despiraliseeritakse, kahe ahela vahelised vesiniksidemed hävivad ensüümide toimel. Ühel DNA ahelal, nagu ka maatriksil, sünteesitakse nukleotiididest RNA koopia vastavalt komplementaarsele põhimõttele. Sõltuvalt DNA piirkonnast sünteesitakse sel viisil ribosomaalseid, transpordi- ja informatsioonilisi RNA-sid.
Pärast mRNA sünteesi lahkub see tuumast ja läheb tsütoplasmasse ribosoomide valgusünteesi kohta.
Saade- polüpeptiidahelate sünteesiprotsess, mis viiakse läbi ribosoomidel, kus mRNA on vahendaja teabe edastamisel valgu primaarse struktuuri kohta.
Valkude biosüntees koosneb reast reaktsioonidest.
1. Aminohapete aktiveerimine ja kodeerimine. tRNA on ristikulehe kujuga, mille keskses ahelas on teatud aminohappe koodile vastav kolmik-antikoodon ja mRNA-l olev koodon. Iga aminohape on ühendatud vastava tRNA-ga, kasutades ATP energiat. Moodustub tRNA-aminohapete kompleks, mis siseneb ribosoomidesse.
2. mRNA-ribosoomi kompleksi moodustumine. Tsütoplasmas olev mRNA on granuleeritud ER-l ühendatud ribosoomidega.
3. Polüpeptiidahela kokkupanek. tRNA aminohapetega, vastavalt antikoodoni ja koodoniga komplementaarsuse põhimõttele, ühinevad mRNA-ga ja sisenevad ribosoomi. Ribosoomi peptiidkeskuses tekib kahe aminohappe vahel peptiidside ja vabanenud tRNA lahkub ribosoomist. Samal ajal liigub mRNA iga kord edasi ühe tripleti, viies sisse uue tRNA – aminohappe ja eemaldades ribosoomist vabanenud tRNA. Kogu protsessi toiteallikaks on ATP. Üks mRNA võib ühineda mitme ribosoomiga, moodustades polüsoomi, kus sünteesitakse samaaegselt palju ühe valgu molekule. Süntees lõpeb, kui mRNA-l algavad mõttetud koodonid (stoppkoodid). Ribosoomid eraldatakse mRNA-st, neilt eemaldatakse polüpeptiidahelad. Kuna kogu sünteesiprotsess toimub granulaarsel endoplasmaatilisel retikulumil, sisenevad tekkivad polüpeptiidahelad EPS tuubulitesse, kus nad omandavad lõpliku struktuuri ja muutuvad valgu molekulideks.
Kõiki sünteesireaktsioone katalüüsivad spetsiaalsed ensüümid, kasutades ATP energiat. Sünteesi kiirus on väga kõrge ja sõltub polüpeptiidi pikkusest. Näiteks Escherichia coli ribosoomis sünteesitakse 300 aminohappest koosnev valk ligikaudu 15-20 sekundiga.
IRNA, tRNA, RRNA – kolme peamise nukleiinhappe – koostoimet ja struktuuri käsitleb selline teadus nagu tsütoloogia. See aitab välja selgitada, milline on transportribonukleiinhappe (tRNA) roll rakkudes. See väga väike, kuid samas vaieldamatult oluline molekul osaleb keha moodustavate valkude ühendamise protsessis.
Mis on tRNA struktuur? Väga huvitav on vaadelda seda ainet "seestpoolt", välja selgitada selle biokeemia ja bioloogiline roll. Ja ka, kuidas on tRNA struktuur ja selle roll valgusünteesis omavahel seotud?
Mis on TRNA, kuidas see on paigutatud?
Transportribonukleiinhape osaleb uute valkude konstrueerimises. Peaaegu 10% kõigist ribonukleiinhapetest on transpordiks. Et oleks selge, millistest keemilistest elementidest molekul moodustub, kirjeldame tRNA sekundaarstruktuuri struktuuri. Sekundaarne struktuur võtab arvesse kõiki peamisi keemilisi sidemeid elementide vahel.
See on makromolekul, mis koosneb polünukleotiidahelast. Selles sisalduvad lämmastikku sisaldavad alused on ühendatud vesiniksidemetega. Nagu DNA-l, on RNA-l 4 lämmastiku alust: adeniin, tsütosiin, guaniin ja uratsiil. Nendes ühendites on adeniin alati seotud uratsiiliga ja guaniin, nagu tavaliselt, tsütosiiniga.
Miks on nukleotiidil eesliide ribo-? Lihtsamalt nimetatakse kõiki lineaarseid polümeere, mille nukleotiidi aluses on pentoosi asemel riboos, ribonukleiinseks. Ja ülekande-RNA on üks kolmest sellise ribonukleiinse polümeeri tüübist.
tRNA struktuur: biokeemia
Vaatame molekuli struktuuri sügavamaid kihte. Nendel nukleotiididel on 3 komponenti:
- Sahharoos, riboos on seotud igat tüüpi RNA-ga.
- Fosforhappe.
- Lämmastiku alused. Need on puriinid ja pürimidiinid.

Lämmastikku sisaldavad alused on omavahel seotud tugevate sidemetega. Aluseid on tavaks jagada puriiniks ja pürimidiiniks.
Puriinid on adeniin ja guaniin. Adeniin vastab kahest omavahel ühendatud tsüklist koosnevale adenüülnukleotiidile. Ja guaniin vastab samale "ühe tsükliga" guaniini nukleotiidile.
Püramidiinid on tsütosiin ja uratsiil. Pürimidiinidel on üks ringstruktuur. RNA-s ei ole tümiini, kuna see on asendatud elemendiga nagu uratsiil. Seda on oluline mõista enne tRNA muude struktuuriliste tunnuste vaatamist.
RNA tüübid
Nagu näete, ei saa tRNA struktuuri lühidalt kirjeldada. Peate süvenema biokeemiasse, et mõista molekuli eesmärki ja selle tegelikku struktuuri. Milliseid teisi ribosomaalseid nukleotiide on teada? Samuti on maatriks- ehk info- ja ribosomaalsed nukleiinhapped. Lühendatult RNA ja RNA. Kõik 3 molekuli teevad rakus üksteisega tihedat koostööd, nii et keha saab õige struktuuriga valgugloobuleid.

Ühe polümeeri tööd on võimatu ette kujutada ilma kahe teise abita. tRNA-de struktuurilised omadused muutuvad arusaadavamaks, kui neid vaadelda koos funktsioonidega, mis on otseselt seotud ribosoomide tööga.
RNA, tRNA, rRNA struktuur on paljuski sarnane. Kõigil on riboosi alus. Nende struktuur ja funktsioonid on aga erinevad.
Nukleiinhapete avastamine
Šveitslane Johann Miescher leidis 1868. aastal raku tuumast makromolekulid, mida hiljem nimetati nukleiinideks. Nimetus "nukleiinid" tuleneb sõnast (tuum) - tuum. Kuigi veidi hiljem leiti, et ainuraksetes olendites, kellel pole tuuma, on need ained samuti olemas. 20. sajandi keskel saadi Nobeli preemia nukleiinhapete sünteesi avastamise eest.
tRNA osaleb valkude sünteesis
Nimetus ise - ülekande-RNA - näitab molekuli põhifunktsiooni. See nukleiinhape "toob" endaga kaasa asendamatu aminohappe, mida ribosomaalne RNA vajab konkreetse valgu valmistamiseks.
tRNA molekulil on vähe funktsioone. Esimene on IRNA koodoni äratundmine, teine funktsioon on ehitusplokkide – aminohapete kohaletoimetamine valgusünteesiks. Veel mõned eksperdid eristavad aktseptori funktsiooni. See tähendab, et aminohapete lisamine kovalentsel põhimõttel. See aitab seda aminohapet "kinnitada" ensüümiga, nagu aminotsiil-tRNA süntaas.
Kuidas on tRNA struktuur seotud selle funktsioonidega? See spetsiaalne ribonukleiinhape on konstrueeritud nii, et selle ühel küljel on lämmastiku alused, mis on alati paarikaupa ühendatud. Need on meile teadaolevad elemendid – A, U, C, G. Täpselt 3 "tähte" ehk lämmastiku alust moodustavad antikoodoni – vastupidise elementide komplekti, mis interakteerub koodoniga vastavalt komplementaarsuse põhimõttele.
See tRNA oluline struktuurne omadus tagab, et matriitsi nukleiinhappe dekodeerimisel ei esine vigu. Sõltub ju aminohapete täpsest järjestusest, kas see valk, mida organism parasjagu vajab, sünteesitakse õigesti.
Struktuursed omadused
Millised on tRNA struktuurilised tunnused ja selle bioloogiline roll? See on väga iidne struktuur. Selle suurus on kuskil 73–93 nukleotiidi. Aine molekulmass on 25 000-30 000.
tRNA sekundaarstruktuuri struktuuri saab lahti võtta, uurides molekuli 5 põhielementi. Niisiis, see nukleiinhape koosneb järgmistest elementidest:
- silmus ensüümiga kokkupuuteks;
- silmus ribosoomiga kokkupuuteks;
- antikoodoni silmus;
- aktseptor vars;
- antikoodon ise.
Ja eraldage sekundaarstruktuuris ka väike muutujasilmus. Igat tüüpi tRNA üks õlg on sama – kahe tsütosiini ja ühe adenosiini jäägi tüvi. Just selles kohas tekib ühendus ühega 20 saadaolevast aminohappest. Iga aminohappe jaoks on ette nähtud eraldi ensüüm - oma aminoatsüül-tRNA.

Kogu teave, mis krüpteerib kõigi nukleiinhapete struktuuri, sisaldub DNA-s endas. Kõigi planeedi elusolendite tRNA struktuur on peaaegu identne. Kahemõõtmelises vaates näeb see välja nagu leht.
Kui aga vaadata mahu järgi, meenutab molekul L-kujulist geomeetrilist struktuuri. Seda peetakse tRNA tertsiaarseks struktuuriks. Kuid õppimise mugavuse huvides on tavaks visuaalselt "lahti keerata". Tertsiaarne struktuur moodustub sekundaarstruktuuri elementide, üksteist täiendavate osade koostoime tulemusena.
tRNA käed või rõngad mängivad olulist rolli. Näiteks on teatud ensüümiga keemilise sideme jaoks vaja ühte kätt.
Nukleotiidi iseloomulik tunnus on tohutu hulga nukleosiidide olemasolu. Neid väiksemaid nukleosiide on rohkem kui 60 tüüpi.
tRNA struktuur ja aminohapete kodeerimine
Teame, et tRNA antikoodon on 3 molekuli pikk. Iga antikoodon vastab konkreetsele, "isiklikule" aminohappele. See aminohape ühendatakse spetsiaalse ensüümi abil tRNA molekuliga. Niipea, kui kaks aminohapet ühinevad, katkevad sidemed tRNA-ga. Kõik keemilised ühendid ja ensüümid on vajalikud kuni vajaliku ajani. Nii on tRNA struktuur ja funktsioonid omavahel seotud.
Kokku on rakus 61 tüüpi selliseid molekule. Matemaatilisi variatsioone võib olla 64. 3 tüüpi tRNA-d aga puuduvad seetõttu, et täpselt nii palju IRNA-s stoppkoodoneid ei ole antikoodoneid.
RNA ja tRNA vastastikmõju
Vaatleme aine interaktsiooni RNA ja RRNA-ga, samuti tRNA struktuurseid iseärasusi. Makromolekuli struktuur ja eesmärk on omavahel seotud.
IRNA struktuur kopeerib teavet DNA eraldi osast. DNA ise on liiga suur molekulide ühendus ja see ei lahku kunagi tuumast. Seetõttu on vaja vahe-RNA-d – informatiivset.

RNA poolt kopeeritud molekulide järjestuse põhjal ehitab ribosoom valku. Ribosoom on eraldiseisev polünukleotiidstruktuur, mille struktuur vajab selgitamist.
Ribosomaalne tRNA: interaktsioon
Ribosomaalne RNA on tohutu organell. Selle molekulmass on 1 000 000 - 1 500 000. Peaaegu 80% RNA koguhulgast moodustavad ribosomaalsed nukleotiidid.

Tundub, et see püüab kinni IRNA ahela ja ootab antikoodoneid, mis toovad endaga kaasa tRNA molekulid. Ribosomaalne RNA koosneb 2 subühikust: väikesed ja suured.
Ribosoomi nimetatakse "tehaseks", kuna selles organellis toimub kogu igapäevaeluks vajalike ainete süntees. See on ka väga iidne rakustruktuur.
Kuidas toimub valkude süntees ribosoomides?
tRNA struktuur ja selle roll valgusünteesis on omavahel seotud. Ribonukleiinhappe ühel küljel paiknev antikoodon sobib oma kujul põhifunktsiooniks - aminohapete toimetamiseks ribosoomi, kus toimub valgu faasiline joondamine. Põhimõtteliselt toimib TRNA vahendajana. Selle ülesandeks on ainult vajalike aminohapete toomine.
Kui RNA ühest osast infot lugeda, liigub ribosoom mööda ahelat edasi. Malli on vaja ainult kodeeritud teabe edastamiseks ühe valgu konfiguratsiooni ja funktsiooni kohta. Järgmisena läheneb ribosoomile teine tRNA oma lämmastikualustega. See dekodeerib ka MRNA järgmise osa.
Dekodeerimine toimub järgmiselt. Lämmastikku sisaldavad alused ühinevad komplementaarsuse põhimõttel samamoodi nagu DNA-s endas. Vastavalt sellele näeb TRNA, kuhu ta peab "silduma" ja millisesse "angaari" aminohappe saata.

Seejärel seotakse ribosoomis sel viisil valitud aminohapped keemiliselt, samm-sammult moodustub uus lineaarne makromolekul, mis pärast sünteesi lõppu keerdub kerakeseks (palliks). Kasutatud tRNA ja RNA, mis on täitnud oma funktsiooni, eemaldatakse valgu "tehasest".
Kui koodoni esimene osa ühineb antikoodoniga, määratakse lugemisraam. Seejärel, kui mingil põhjusel toimub kaadri nihe, lükatakse mõni valgu märk tagasi. Ribosoom ei saa sellesse protsessi sekkuda ja probleemi lahendada. Alles pärast protsessi lõppu ühendatakse 2 rRNA subühikut uuesti. Keskmiselt on iga 10 4 aminohappe kohta 1 viga. Iga 25 juba kokkupandud valgu kohta ilmneb kindlasti vähemalt 1 replikatsiooniviga.
tRNA kui reliktmolekulid
Kuna tRNA võis eksisteerida ajal, mil elu Maal sündis, nimetatakse seda reliikvia molekuliks. Arvatakse, et RNA on esimene struktuur, mis eksisteeris enne DNA-d ja seejärel arenes. RNA maailma hüpotees – sõnastas 1986. aastal laureaat Walter Gilbert. Seda on aga endiselt raske tõestada. Teooriat kaitsevad ilmsed faktid – tRNA molekulid suudavad salvestada infoplokke ja seda infot kuidagi rakendada ehk tööd teha.
Kuid teooria vastased väidavad, et aine lühike eluiga ei saa garanteerida, et tRNA on hea igasuguse bioloogilise teabe kandja. Need nukleotiidid lagunevad kiiresti. tRNA eluiga inimese rakkudes ulatub mõnest minutist mitme tunnini. Mõned liigid võivad kesta kuni päeva. Ja kui me räägime samadest nukleotiididest bakterites, siis on terminid palju lühemad - kuni mitu tundi. Lisaks on tRNA struktuur ja funktsioonid liiga keerulised, et molekulist saaks Maa biosfääri esmane element.
Kõigil tRNA-del on ühised jooned nii nende primaarstruktuuris kui ka viisis, kuidas polünukleotiidahelat nukleotiidjääkide aluste vastastikmõju tõttu sekundaarseks struktuuriks voltida.
tRNA esmane struktuur
tRNA-d on suhteliselt väikesed molekulid, nende ahela pikkus varieerub 74 kuni 95 nukleotiidi jäägi vahel. Kõikidel tRNA-del on sama 3'-ots, mis on üles ehitatud kahest tsütosiini ja ühest adenosiini jäägist (CCA-ots).See on 3'-otsa adenosiin, mis seondub aminohappejäägiga aminoatsüül-tRNA moodustumisel. CCA ots on paljude tRNA-de külge kinnitatud spetsiaalse ensüümi abil. Aminohappekoodoniga (antikoodon) komplementaarne nukleotiidi kolmik paikneb ligikaudu tRNA ahela keskel. Peaaegu igat tüüpi tRNA-s leidub järjestuse teatud positsioonides samu (konservatiivseid) nukleotiidijääke. Mõned positsioonid võivad sisaldada kas ainult puriini või ainult pürimidiini aluseid (neid nimetatakse poolkonservatiivseteks jääkideks).
Kõiki tRNA molekule iseloomustab suur hulk (kuni 25% kõigist jääkidest) mitmesuguste modifitseeritud nukleosiidide, mida sageli nimetatakse väikesteks, olemasolu. Need moodustuvad molekulide erinevates kohtades, paljudel juhtudel täpselt määratletud, tavaliste nukleosiidijääkide modifitseerimise tulemusena spetsiaalsete ensüümide abil.
tRNA sekundaarne struktuur
ahela voltimine sekundaarstruktuuriks toimub keti osade vastastikuse komplementaarsuse tõttu. Kolm keti fragmenti täiendavad üksteist, kui need kokku voltida, moodustades juuksenõelastruktuure. Lisaks täiendab 5-tolline ots keti 3-tollise otsa lähedal asuvat kohta oma antiparalleelse paigutusega; nad moodustavad nn aktseptortüve. Tulemuseks on struktuur, mida iseloomustab nelja varre ja kolme silmuse olemasolu, mida nimetatakse "ristikleheks". Aasaga vars moodustab oksa. Allosas on antikoodoni haru, mis sisaldab oma silmuse osana antikoodoni kolmikut. Sellest vasakul ja paremal on D- ja T-harud, mis on nimetatud vastavalt ebatavaliste konserveerunud dihüdrouridiini (D) ja tümidiini (T) nukleosiidide olemasolu kohta nende ahelates. Kõigi uuritud tRNA-de nukleotiidjärjestusi saab voltida sarnasteks struktuurideks. Lisaks kolmele ristikulehe ahelale isoleeritakse tRNA struktuuris ka täiendav ehk muutuv silmus (V-silmus). Selle suurus on erinevates tRNA-des järsult erinev, varieerudes 4–21 nukleotiidi ja viimastel andmetel kuni 24 nukleotiidini.
tRNA ruumiline (tertsiaarne) struktuur
Sekundaarse struktuuri elementide koosmõjul tekib tertsiaarne struktuur, mida nimetatakse L-vormiks sarnasuse tõttu ladina tähega L (joon. 2 ja 3). Aluse virnastamise kaudu moodustavad aktseptori vars ja ristikulehe T-tüvi ühe pideva topeltheeliksi ning ülejäänud kaks vart moodustavad antikoodoni ja D-tüve teise pideva kaksikheeliksi. Sel juhul osutuvad D- ja T-aasad tihedaks ja kinnitatakse kokku, moodustades täiendavaid, sageli ebatavalisi aluspaare. Reeglina osalevad nende paaride moodustamisel konservatiivsed või poolkonservatiivsed jäägid. Sarnased tertsiaarsed interaktsioonid hoiavad koos ka mõningaid teisi L-struktuuri osi
Ribosomaalne RNA
Ribosomaalsed ribonukleiinhapped (rRNA) on mitmed RNA molekulid, mis moodustavad ribosoomi aluse. rRNA põhiülesanne on translatsiooniprotsessi elluviimine – mRNA-st informatsiooni lugemine adapter-tRNA molekulide abil ja peptiidsidemete moodustumise katalüüsimine tRNA-ga seotud aminohapete vahel. Ribosomaalne RNA moodustab ligikaudu 80% kogu raku RNA-st. Seda kodeerivad geenid, mis asuvad mitme kromosoomi DNA-s, mis paiknevad tuuma piirkonnas, mida tuntakse nukleolaarse organisaatorina.
Alusjärjestus rRNA-s on sarnane kõikides organismides, alates bakteritest kuni loomadeni. rRNA-d leidub tsütoplasmas, kus see on seotud valgu molekulidega, moodustades nendega rakuorganellid, mida nimetatakse ribosoomideks. Valkude süntees toimub ribosoomidel. Siin transleeritakse mRNA-s sisalduv "kood" polüpeptiidahela aminohappejärjestuseks.
RNA ülekandmine
Transfer RNA, tRNA - ribonukleiinhape, mille ülesanne on aminohapete transportimine valgusünteesi kohta. tRNA-d osalevad otseselt ka polüpeptiidahela kasvus, liitudes - olles kompleksis aminohappega - mRNA koodoniga ja tagades kompleksi uue peptiidsideme moodustamiseks vajaliku konformatsiooni.
Igal aminohappel on oma tRNA.
tRNA on üheahelaline RNA, kuid funktsionaalsel kujul on tal "ristiklehe" konformatsioon. Sellel on neli põhiosa, mis täidavad erinevaid funktsioone. Aktseptori "vars" moodustub kahest komplementaarsest ühendatud terminaalsest tRNA osast. See koosneb seitsmest aluspaarist. Selle varre 3" ots on mõnevõrra pikem ja moodustab üheahelalise piirkonna, mis lõpeb vaba OH-rühmaga CCA järjestuses. Sellele otsale on kinnitatud transporditav aminohape. Ülejäänud kolm haru on komplementaarselt paaritud nukleotiidjärjestused, mis lõpevad paaritute ahelat moodustavates piirkondades. Nende harude keskmine – antikoodon – koosneb viiest nukleotiidipaarist ja sisaldab oma ahela keskel antikoodonit.Antikoodon on kolm nukleotiidi, mis on komplementaarsed mRNA koodoniga, mis kodeerib aminohapet transporditakse selle tRNA abil peptiidide sünteesikohta.
Aktseptori ja antikoodoni harude vahel on kaks külgharu. Oma ahelates sisaldavad nad modifitseeritud aluseid - dihüdrouridiini (D-silmus) ja T?C tripletti, kus? - pseudouriaiin (T? C-silmus). Aitikodoni ja T?C harude vahel on täiendav silmus, mis sisaldab 3-5 kuni 13-21 nukleotiidi.
Aminohape on kovalentselt kinnitatud molekuli 3' otsa ensüümi aminoatsüül-tRNA süntetaas, mis on spetsiifiline iga tRNA tüübi jaoks.
tRNA toimib vahemolekulina mRNA kolmikkoodoni ja polüpeptiidahela aminohappejärjestuse vahel. tRNA moodustab ligikaudu 15% kogu raku RNA-st; neil RNA-del on lühim polünukleotiidahel - see sisaldab keskmiselt 80 nukleotiidi. Iga rakk sisaldab rohkem kui 20 erinevat tRNA molekuli. Kõigil tRNA molekulidel on sarnane põhistruktuur. tRNA molekuli 5'-otsas on alati guaniin ja 3'-otsas - CCA alusjärjestus.
Nukleotiidjärjestus ülejäänud molekulis varieerub ja võib sisaldada "ebatavalisi" aluseid, nagu inosiin ja pseudouratsiil.
Antikoodoni kolmiku alusjärjestus vastab rangelt aminohappele, mida antud tRNA molekul kannab.
Riis. 3.
Iga aminohape kinnitub ensüümi aminoatsüül-tRNA süntaasi abil ühele oma spetsiifilistest tRNA-dest. Tulemuseks on animatsüül-tRNA kompleks, tuntud kui animoatsüül-tRNA, milles CCA-tripleti terminaalse nukleotiidi A ja aminohappe vaheliste sidemete energia on piisav, et võimaldada edasist sidet külgneva aminohappega. Seega sünteesitakse polüpeptiidahel.
Üks tRNA tunnuseid on ebatavaliste aluste olemasolu selles, mis tekivad keemilise modifitseerimise tulemusena pärast normaalse aluse kaasamist polünukleotiidahelasse. Need muudetud alused määravad tRNA-de suure struktuurilise mitmekesisuse nende struktuuri üldplaanis. Suurimat huvi pakuvad antikoodoni moodustavate aluste modifikatsioonid, mis mõjutavad selle interaktsiooni spetsiifilisust koodoniga. Näiteks ebatüüpiline alusinosiin, mõnikord tRNA antikoodoni 1. positsioonis, on võimeline komplementaarselt kombineeruma mRNA koodoni kolme erineva kolmanda alusega - U, C ja A. Kuna geneetilise koodi üheks tunnuseks on selle degeneratsiooni korral on paljud aminohapped krüpteeritud mitme koodoniga, mis reeglina erinevad oma kolmanda aluse poolest. Modifitseeritud antikoodoni aluse mittespetsiifilise seondumise tõttu tunneb üks tRNA ära mitu sünonüümset koodonit.