Metoder for cytologimateriale for forberedelse til eksamen. Cytologi
Mishnina Lidia Alexandrovna
biologilærer
MBOU ungdomsskole nr. 3 Akbulak landsby
Klasse 11
Forberedelse til eksamen: løse problemer i cytologi
I retningslinjene for forbedring av undervisningen i biologi, utviklet på grunnlag av en analyse av vanskene til nyutdannede i USE i 2014, skrev forfatterne G.S. Kalinova, R.A. Petrosova, er det et lavt nivå av utførelse av oppgaver for å bestemme antall kromosomer og DNA i forskjellige faser av mitose eller meiose.
Oppgavene er faktisk ikke så vanskelige at de skaper alvorlige vanskeligheter. Hva bør tas i betraktning når du forbereder nyutdannede på dette problemet?
Løsningen av cytologiske problemer innebærer kunnskap ikke bare om spørsmålene om mitose og meiose, deres faser og hendelser som oppstår i dem, men også den obligatoriske besittelsen av informasjon om strukturen og funksjonene til kromosomer, mengden genetisk materiale i cellen.
Derfor begynner vi forberedelsen med å gjenta materialet om kromosomer. Vi fokuserer på det faktum at kromosomer er nukleoproteinstrukturer i kjernen til en eukaryot celle.
Omtrent 99% av hele cellens DNA er konsentrert i dem, resten av DNA er lokalisert i andre cellulære organeller, og bestemmer cytoplasmatisk arv. DNA i eukaryote kromosomer er i kompleks med hovedproteinene - histoner og ikke-histonproteiner, som gir kompleks pakking av DNA i kromosomer og regulerer dets evne til å syntetisere ribonukleinsyrer (RNA) - transkripsjon.
Utseendet til kromosomer endres betydelig på forskjellige stadier av cellesyklusen, og som kompakte formasjoner med en karakteristisk morfologi, er kromosomene klart å skille i et lysmikroskop bare i celledelingsperioden.
På metafasestadiet av mitose og meiose består kromosomene av to langsgående kopier, som kalles søsterkromatider og som dannes under DNA-replikasjon i S-perioden av interfase. I metafasekromosomer er søsterkromatider forbundet ved den primære innsnevringen, kalt sentromeren. Sentromeren er ansvarlig for å skille søsterkromatider til datterceller under deling.
Det komplette settet med kromosomer i en celle, karakteristisk for en gitt organisme, kalles en karyotype. I en hvilken som helst celle i kroppen til de fleste dyr og planter er hvert kromosom representert to ganger: en av dem ble mottatt fra faren, den andre fra moren under sammensmeltingen av kjernene til kjønnsceller under befruktning. Slike kromosomer kalles homologe, settet med homologe kromosomer kalles diploide.
Nå kan du gjenta materialet på celledeling.
Av interfasehendelsene vurderer vi bare den syntetiske perioden, for ikke å spre oppmerksomheten til skolebarn, men bare fokusere på kromosomenes oppførsel.
Husk: i den syntetiske (S) perioden dobles arvematerialet ved DNA-replikasjon. Det skjer på en semi-konservativ måte, når den doble helixen til DNA-molekylet divergerer i to tråder og en komplementær tråd syntetiseres på hver av dem.
Som et resultat dannes det to identiske DNA-dobbelthelixer, som hver består av en ny og en gammel DNA-streng. Mengden arvelig materiale dobles, men antallet kromosomer forblir det samme - kromosomet blir to-kromatid (2n4c).
Vurder oppførselen til kromosomer under mitose:
- I profase, metafase - 2p 4s - siden celledeling ikke forekommer;
- I anafase skiller kromatider seg, antall kromosomer dobles (kromatider blir uavhengige kromosomer, men så langt er de alle i én celle) 4n 4с;
- i telofase 2p2c (enkeltkromatidkromosomer forblir i cellene).
Vi gjentar meiose:
- I profase 1, metafase 1, anafase 1 - 2p 4s - siden celledeling ikke forekommer;
- i telofase - p2c gjenstår, siden etter divergensen av homologe kromosomer, forblir et haploid sett i cellene, men kromosomene er to-kromatider;
- I profase 2, metafase 2 samt telofase 1 - p2s;
- Vær spesielt oppmerksom på anafase 2, siden etter kromatidseparasjon øker antall kromosomer med 2 ganger (kromatider blir uavhengige kromosomer, men så langt er de alle i en celle) 2n 2с;
- i telofase 2 - ps (enkeltkromatidkromosomer forblir i cellene.
Først nå, når barna er teoretisk forberedt, kan vi gå videre til å løse problemer.
En typisk feil i forberedelsen av nyutdannede: vi prøver å umiddelbart løse problemer uten å gjenta materialet. Hva skjer: barna og læreren bestemmer, men avgjørelsen er på nivået utenat utenat, uten forståelse. Derfor, når de får en lignende oppgave på eksamen, takler de det ikke. Jeg gjentar: det var ingen forståelse for å løse problemer.
La oss gå videre til praksis.
Vi bruker et utvalg oppgaver fra nettstedet "Jeg skal løse eksamen" av Dmitry Gushchin. Det som er attraktivt med denne ressursen er at det praktisk talt ikke er noen feil, svarstandardene er velskrevne.
La oss analysere problem C 6 nr. 12018.
Kromosomsettet av somatiske hveteceller er 28.
Bestem kromosomsettet og antall DNA-molekyler i en av cellene i eggløsningen før meiose, i anafasen til meiose 1 og i anafasen til meiose 2. Forklar hvilke prosesser som skjer i disse periodene og hvordan de påvirker endringen i antall DNA og kromosomer.
Responselementer:
Eggløsningsceller inneholder et diploid sett med kromosomer - 28 (2n2c).
Før meiose - (2n4c) 28 xp, 56 DNA
I meiotisk anafase 1: (2n4c = n2c+n2c) - 28 xp, 56 DNA.
Meiose 2 legges inn av 2 datterceller med et haploid sett av kromosomer (n2c) - 14 kromosomer, 28DNA.
I meiose anafase 2: (2n2с= nc+nc) - 28 kromosomer, 28DNA
Oppgaven er vanskelig, hvordan hjelpe kandidaten til å forstå løsningen.
Ett av alternativene: vi tegner fasene til meiose og viser alle manipulasjoner med kromosomer.
Handlingsalgoritme:
- Les oppgaven nøye, definer oppgaven, skriv ned fasene der du må angi mengden genetisk materiale
a) Før starten av meiose
b) I anafasen av meiose 1
c) I anafasen av meiose 2
- Lag tegninger for hver utpekt fase av meiose og forklar hva du har gjort.
Jeg presiserer: vi bruker ikke tegninger, men vi lager dem selv. Denne operasjonen fungerer på forståelse ( selv om vi taper i estetikk, vinner vi som et resultat!)
1. Før meiose
Jeg forklarer: meiose innledes med interfase, DNA-dobling skjer i interfase, derfor er antall kromosomer 2p, antall DNA er 4c.
2. I anafasen av meiose 1
Jeg forklarer: i anafasen til meiose 1 divergerer kromosomene til polene, dvs. av hvert par homologe kromosomer kommer bare ett inn i dattercellen. Kromosomsettet blir haploid, men hvert kromosom består av to kromatider. Siden celledeling ennå ikke har funnet sted og alle kromosomene er i én celle, kan kromosomformelen skrives som: 2n4c (n2c + n2c) 28 хр, 56 DNA (14хр 28 DNA + 14хр28DNA)
3) I anafasen av meiose 2
Meiotisk anafase 2 oppstår etter den første (reduksjons)delingen. Et sett med kromosomer i en p2c-celle. I anafasen av meiose deler de 2 sentromerene som forbinder søsterkromatidene seg og kromatidene, som i mitose, blir uavhengige kromosomer. Antall kromosomer øker og blir lik 2n2c. Og igjen, siden celledeling ennå ikke har skjedd og alle kromosomene er i en celle, kan kromosomsettet skrives som følger: 2n2c (nc + nc) 28 хр, 28 DNA (14хр 14 DNA + 14хр14DNA).
- Skriv ned svaret. (vi har det over)
Jeg oppsummerer: Å løse problemer av denne typen krever ikke jakten på kvantitet, her er det viktig å oppnå en forståelse av logikken i løsningen og kunnskap om kromosomenes oppførsel i hver fase av delingen.
Ressurser brukt:
- FIPI "Metodologiske anbefalinger om noen aspekter ved å forbedre undervisningen i biologi" utg. G.S. Kalinova, R.A. Petrosov. Moskva, 2014
- Biologi. Generelle mønstre Grad 10: en lærebok for utdanningsinstitusjoner / V.B. Zakharov, S.G. Mamontov, N.I. Sonin - Moskva: Drofa Publishing House, 2011.
- Jeg skal løse eksamen. http://bio.reshuege.ru/
Kjære lesere! Hvis du velger USE som avsluttende eller opptaksprøve i biologi, må du kjenne til og forstå kravene for å bestå denne eksamen, arten av spørsmålene og oppgavene som finnes i eksamensoppgavene. For å hjelpe søkere vil EKSMO forlag gi ut boken «Biologi. Samling av oppgaver for forberedelse til eksamen. Denne boken er en opplæringsmanual, og det er grunnen til at materialet som er inkludert i den overskrider skolekravene. Men for de videregående studenter som bestemmer seg for å gå inn på høyere utdanningsinstitusjoner ved fakulteter der biologi tas, vil denne tilnærmingen være nyttig.
I avisen vår publiserer vi kun oppgavene i del C for hver seksjon. De er fullstendig oppdatert både i innhold og i presentasjonsstrukturen. Siden denne håndboken er fokusert på eksamenene i studieåret 2009/2010, bestemte vi oss for å gi alternativer for del C-oppgaver i et mye større volum enn det som ble gjort tidligere år.
Du tilbys omtrentlige alternativer for spørsmål og oppgaver med forskjellige kompleksitetsnivåer med et annet antall elementer av riktig svar. Dette gjøres for at du på eksamen skal ha et tilstrekkelig stort utvalg av mulige riktige svar på et spesifikt spørsmål. I tillegg er spørsmålene og oppgavene i del C strukturert som følger: ett spørsmål og elementene i det riktige svaret på det er gitt, og deretter tilbys varianter av dette spørsmålet for uavhengig refleksjon. Svarene på disse alternativene bør du få tak i selv, ved å bruke både kunnskapen du har fått fra å studere materialet og kunnskapen du får ved å lese svarene på hovedspørsmålet. Alle spørsmål skal besvares skriftlig.
En vesentlig del av oppgavene i del C er oppgaver i tegninger. Lignende dem var allerede i eksamensoppgavene fra 2008. I denne håndboken er settet deres noe utvidet.
Vi håper at denne læreboken vil hjelpe elever på videregående skole til ikke bare å forberede seg til eksamen, men også gi en mulighet for de som ønsker å lære det grunnleggende om biologi i de resterende to studieårene på 10.-11. trinn.
Generell biologi (del C)
Oppgavene til denne delen er delt inn i seksjoner: cytologi, genetikk, evolusjonsteori, økologi. Hver seksjon tilbyr oppgaver for alle nivåer av eksamen. En slik konstruksjon av den generelle biologiske delen av manualen vil tillate deg å mer fullstendig og systematisk forberede deg til eksamen, fordi. Del C inkluderer, i en generalisert form, nesten alt materialet i del A og B.
Oppgaver til gruppe C1 (avansert nivå)
Alle oppgaver i gruppe C skal besvares skriftlig med forklaringer.
Spørsmål om cytologi
Svaret på dette spørsmålet bør være kort, men presist. Ordene "organisasjonsnivåer" og "vitenskapelige grunnlag" er de viktigste i denne saken. Organisasjonsnivået er måten og eksistensformen for levende systemer. For eksempel inkluderer det cellulære organisasjonsnivået celler. Derfor er det nødvendig å finne ut hva som er vanlig, noe som gjorde det mulig å skille organisasjonsnivåene. Et slikt fellestrekk er den systematiske organiseringen av levende kropper og deres gradvise komplikasjon (hierarki).
Elementer av riktig svar
Det vitenskapelige grunnlaget for å dele inn levende systemer i nivåer er følgende bestemmelser.
1. Levende systemer blir mer komplekse etter hvert som de utvikler seg: celle - vev - organisme - populasjon - arter osv.
2. Hvert mer høyt organisert levende system inkluderer de tidligere systemene. Vev består av celler, organer består av vev, organismer består av organer, og så videre.
Svar på følgende spørsmål selv
Hvilke felles egenskaper har alle nivåer i livsorganisasjonen?
Hva er likhetene og forskjellene mellom celle- og befolkningsnivået i livet?
Bevis at alle egenskapene til levende systemer er manifestert på cellenivå.
Elementer av riktig svar
1. Det er mulig å anvende påvirkninger på modellen som ikke er anvendelige for levende kropper.
2. Modellering lar deg endre alle egenskaper ved objektet.
Svar selv
Hvordan vil du forklare uttalelsen til I.P. Pavlova «Observasjon samler det naturen tilbyr den, mens opplevelsen tar fra naturen det den vil ha»?
Gi to eksempler på bruk av den eksperimentelle metoden i cytologi.
Hvilke forskningsmetoder kan brukes for å skille ulike cellulære strukturer?
Elementer av riktig svar
1. Polariteten til et vannmolekyl bestemmer dets evne til å løse opp andre hydrofile stoffer.
2. Vannmolekylers evne til å danne og bryte hydrogenbindinger mellom dem gir vann varmekapasitet og termisk ledningsevne, overgangen fra en aggregeringstilstand til andre.
3. Den lille størrelsen på molekylene sikrer deres evne til å penetrere mellom molekylene til andre stoffer.
Svar selv
Hva vil skje med cellen hvis konsentrasjonen av salter i den er høyere enn utenfor cellen?
Hvorfor krymper ikke cellene og brister av hevelse i fysiologisk saltvann?
Elementer av riktig svar
1. Forskere har funnet ut at et proteinmolekyl har primære, sekundære, tertiære og kvartære strukturer.
2. Forskere har funnet ut at et proteinmolekyl består av mange forskjellige aminosyrer knyttet sammen med peptidbindinger.
3. Forskere har etablert sekvensen av aminosyrerester i ribonukleasemolekylet, dvs. dens primære struktur.
Svar selv
Hvilke kjemiske bindinger er involvert i dannelsen av et proteinmolekyl?
Hvilke faktorer kan føre til proteindenaturering?
Hva kjennetegner strukturen og funksjonene til enzymer?
I hvilke prosesser manifesteres beskyttelsesfunksjonene til proteiner?
Elementer av riktig svar
1. Disse organiske forbindelsene utfører en bygningsmessig (strukturell) funksjon.
2. Disse organiske forbindelsene utfører en energifunksjon.
Svar selv
Hvorfor foreskrives celluloserik mat for å normalisere tarmfunksjonen?
Hva er byggefunksjonen til karbohydrater?
Elementer av riktig svar
1. DNA er bygget på prinsippet om en dobbel helix i samsvar med regelen om komplementaritet.
2. DNA består av repeterende elementer - 4 typer nukleotider. Ulik sekvens av nukleotider koder for forskjellig informasjon.
3. DNA-molekylet er i stand til selvreproduksjon, og derfor til å kopiere informasjon og dens overføring.
Svar selv
Hvilke fakta beviser individualiteten til et individs DNA?
Hva betyr begrepet "den genetiske kodens universalitet"; hvilke fakta bekrefter denne universaliteten?
Hva er den vitenskapelige verdien til D. Watson og F. Crick?
Elementer av riktig svar
1. Forskjeller i navnene på DNA og RNA forklares av sammensetningen av deres nukleotider: i DNA-nukleotider er karbohydratet deoksyribose, og i RNA ribose.
2. Forskjeller i navn på RNA-typer (informasjons-, transport-, ribosomalt) er knyttet til funksjonene de utfører.
Svar selv
Hvilke to forhold må være konstante for at bindingene mellom to komplementære DNA-tråder ikke skal bryte spontant?
Hvordan skiller DNA og RNA seg i struktur?
Hvilke andre forbindelser inneholder nukleotider, og hva vet du om dem?
Elementer av riktig svar
1. Celleteori etablerte den strukturelle og funksjonelle enheten til de levende.
2. Celleteorien etablerte enheten for reproduksjon og utvikling av de levende.
3. Celleteori bekreftet den vanlige strukturen og opprinnelsen til levende systemer.
Svar selv
Hvorfor, til tross for de åpenbare forskjellene i strukturen og funksjonene til cellene i forskjellige vev, snakker de om enheten i den cellulære strukturen til de levende?
Hva er hovedfunnene i biologien som gjorde det mulig å formulere celleteorien.
Elementer av riktig svar
1. Stoffer kommer inn i cellen ved diffusjon.
2. Stoffer kommer inn i cellen på grunn av aktiv transport.
3. Stoffer kommer inn i cellen ved pinocytose og fagocytose.
Svar selv
Hva er forskjellen mellom aktiv transport av stoffer over cellemembranen og passiv transport?
Hvilke stoffer fjernes fra cellen og hvordan?
Elementer av riktig svar
1. Hos prokaryoter mangler cellen en kjerne, mitokondrier, Golgi-apparatet og endoplasmatisk retikulum.
2. Prokaryoter har ikke ekte seksuell reproduksjon.
Svar selv
Hvorfor er modne erytrocytter eller blodplater ikke klassifisert som prokaryote celler, til tross for fravær av kjerner i dem?
Hvorfor klassifiseres ikke virus som uavhengige organismer?
Hvorfor er eukaryote organismer mer forskjellige i struktur og kompleksitet?
Elementer av riktig svar
1. Ved hjelp av kromosomsettet til et dyr kan du bestemme typen.
2. Ved kromosomsettet til et dyr kan du bestemme kjønnet.
3. I henhold til kromosomsettet til et dyr er det mulig å bestemme tilstedeværelsen eller fraværet av arvelige sykdommer.
Svar selv
Har hver celle i en flercellet organisme kromosomer? Bevis svaret ditt med eksempler.
Hvordan og når kan du se kromosomer i en celle?
Elementer av riktig svar
De strukturelle elementene i Golgi-komplekset er:
1) tubuli;
2) hulrom;
3) bobler.
Svar selv
Hva er strukturen til en kloroplast?
Hva er strukturen til et mitokondrie?
Hva må inneholdes i mitokondrier slik at de kan syntetisere proteiner?
Bevis at både mitokondrier og kloroplaster kan formere seg.
Elementer av riktig svar
Legg merke til forskjellene i:
1) stoffskiftets natur;
2) livsvilkår;
3) reproduksjon.
Svar selv
Hvordan vil transplantasjon av en kjerne fra en annen organisme påvirke en encellet organisme?
Elementer av riktig svar
1. Tilstedeværelsen av en dobbel membran med karakteristiske kjerneporer, som sikrer forbindelsen av kjernen med cytoplasma.
2. Tilstedeværelsen av nukleoler, der RNA syntetiseres og ribosomer dannes.
3. Tilstedeværelsen av kromosomer, som er cellens arvelige apparat og sørger for nukleær deling.
Svar selv
Hvilke celler inneholder ikke kjerner?
Hvorfor reproduserer ikke-nukleære prokaryote celler, men ikke-nukleære eukaryote celler gjør det ikke?
Elementer av riktig svar
1. De fleste celler er like i grunnleggende strukturelle elementer, vitale egenskaper og delingsprosessen.
2. Celler skiller seg fra hverandre i nærvær av organeller, spesialisering i funksjonene som utføres, og intensiteten av metabolisme.
Svar selv
Gi eksempler på samsvaret mellom strukturen til en celle og dens funksjon.
Gi eksempler på celler med ulike nivåer av metabolsk intensitet.
Elementer av riktig svar
1. Som et resultat av syntese dannes det mer komplekse stoffer enn de som har reagert; reaksjonen fortsetter med absorpsjon av energi.
2. Under forfallet dannes det enklere stoffer enn de som har reagert; Reaksjonen fortsetter med frigjøring av energi.
Svar selv
Hva er funksjonene til enzymer i metabolske reaksjoner?
Hvorfor er mer enn 1000 enzymer involvert i biokjemiske reaksjoner?
17. Hvilke typer energi blir lysenergi til under fotosyntesen og hvor finner denne transformasjonen sted? |
Elementer av riktig svar
1. Lysenergi omdannes til kjemisk og termisk energi.
2. Alle transformasjoner skjer i thylakoidene til grankloroplastene og i deres matrise (i planter); i andre fotosyntetiske pigmenter (i bakterier).
Svar selv
Hva skjer i lysfasen av fotosyntesen?
Hva skjer i den mørke fasen av fotosyntesen?
Hvorfor er det vanskelig å eksperimentelt oppdage prosessen med planterespirasjon på dagtid?
Elementer av riktig svar
1. Koden "triplett" betyr at hver av aminosyrene er kodet av tre nukleotider.
2. Koden er "entydig" - hver triplett (kodon) koder kun for én aminosyre.
3. Koden "degenerert" betyr at hver aminosyre kan kodes av mer enn ett kodon.
Svar selv
Hvorfor trenger vi "tegningstegn" mellom gener og hvorfor er de ikke inne i gener?
Hva betyr konseptet "universaliteten til DNA-koden"?
Hva er den biologiske betydningen av transkripsjon?
Elementer av riktig svar
1. Eksempler på organismer hvor generasjonsveksling skjer kan være moser, bregner, maneter og andre.
2. Hos planter endres gametofytten og sporofytten. Maneter har vekslende polypp- og medusa-stadier.
Svar selv
Hva er hovedforskjellene mellom mitose og meiose?
Hva er forskjellen mellom begrepene "cellesyklus" og "mitose"?
Elementer av riktig svar
1. Isolerte kroppsceller som lever i et kunstig miljø kalles cellekultur (eller cellekultur).
2. Cellekulturer brukes til å skaffe antistoffer, medikamenter, samt for å diagnostisere sykdommer.
Elementer av riktig svar
1. Interfase er nødvendig for lagring av stoffer og energi som forberedelse til mitose.
2. I interfasen dobles arvematerialet, noe som deretter sikrer ensartet fordeling blant dattercellene.
Svar selv
Er kjønnscellene produsert av en organisme like eller forskjellige i deres genetiske sammensetning? Kom med bevis.
Hvilke organismer har en evolusjonær fordel - haploid eller diploid? Kom med bevis.
Nivå C2 oppgaver
Elementer av riktig svar
Det ble gjort feil i setningene 2, 3, 5.
I setning 2, legg merke til et av ikke-makroelementene.
I setning 3 er ett av de oppførte elementene feilaktig tilordnet mikroelementer.
I setning 5 er elementet som utfører den navngitte funksjonen feilaktig angitt.
2. Finn feil i den gitte teksten. Angi antall setninger der feil ble gjort, forklar dem. 1. Proteiner er uregelmessige biopolymerer hvis monomerer er nukleotider. 2. Restene av monomerene er forbundet med peptidbindinger. 3. Sekvensen av monomerer støttet av disse bindingene danner den primære strukturen til proteinmolekylet. 4. Den neste strukturen er sekundær, støttet av svake hydrofobe kjemiske bindinger. 5. Den tertiære strukturen til et protein er et vridd molekyl i form av en kule (kule). 6. Denne strukturen er støttet av hydrogenbindinger. |
Elementer av riktig svar
Det ble gjort feil i setningene 1, 4, 6.
I setning 1 er monomerene til proteinmolekylet feil angitt.
Setning 4 indikerer feil de kjemiske bindingene som støtter den sekundære strukturen til proteinet.
Setning 6 indikerer feil de kjemiske bindingene som støtter tertiærstrukturen til proteinet.
LØSE PROBLEMER I CYTOLOGI
Læremiddel
Ny Adelakovo 2014
Satt sammen av: biologilærer L.I. Denisova
Løse problemer i cytologi. For elever i klasse 9-11 / GBOU videregående skole v. Novoe Adelakovo "; komp. L.I.Denisova. – Nye Adelakovo, 2014.
Læremidlet er satt sammen med det formål å bruke biologilærere og studenter som forberedelse til den endelige sertifiseringen, samt til biologi-olympiader. Manualen inneholder kort teoretisk stoff om hver type problemstilling og eksempler på problemløsning. Designet for studenter i klasse 9-11 ved utdanningsinstitusjoner og biologilærere.
Introduksjon……………………………………………………………..
2.1 Oppgaver relatert til å bestemme prosentandelen av nukleotider i DNA
2.2. R
2.3. Oppgaver for konstruksjon av et i-RNA-molekyl, t-RNA-antikodoner og aminosyresekvenser i en polypeptidkjede. Arbeid
Litteratur.
Introduksjon.
Løsningen av problemer i cytologi er inkludert i KIM i biologi i Unified State Examination (39 oppgave). Gjennomføringen av denne oppgaven gir et detaljert svar og er rettet mot å teste ferdigheter.
- anvende kunnskap i en ny situasjon;
- etablere årsakssammenhenger;
- analysere, systematisere og integrere kunnskap;
- oppsummere og formulere konklusjoner.
I følge resultatene av analysen av resultatene fra USE i biologi, er følgende blant de dårlig utformede kunnskapene og ferdighetene til deltakerne i eksamen:
- bestemmelse av kromosomsettet av celler i planteutviklingssykluser;
- bestemmelse av antall kromosomer og DNA i celler i ulike faser av mitose og meiose;
- forklaring og begrunnelse av resultatet.
Når man studerer biologi på et grunnleggende nivå, er det ikke nok tid til å trene på å løse problemer i cytologi. Etter å ha gått gjennom relevante emner, uten konstant repetisjon av den praktiske utviklingen av problemløsning, blir ferdighetene raskt glemt. Studentene kan alltid ha denne håndboken for hånden for å huske forløpet med å løse typiske problemer. Spesielt på landsbygda har ikke alle gratis tilgang til Internett-ressurser.
Oppgaver i cytologi som finnes på eksamen kan deles inn i flere hovedtyper. Denne håndboken tilbyr løsninger på ulike typer problemer og gir eksempler for selvstendig arbeid. Vedlegget inneholder en tabell over den genetiske koden som brukes i løsningen.
Dette læremiddelet er satt sammen for å hjelpe biologilærere som studerer i klasse 9-11 på ungdomsskoler.
1.1 Nøkkelpunkter å huske på når man løser problemer i cytologi.
- Hver aminosyre leveres til ribosomene til ett tRNA, derforantall aminosyrer i et protein er lik antall tRNA-molekylerinvolvert i proteinsyntese;
- hver aminosyre er kodet av tre nukleotider (en triplett eller kodon), så antallet kodende nukleotider er alltid tre ganger større, ogantall tripletter (kodoner) er lik antall aminosyrer i proteinet;
- hvert tRNA har et antikodon som er komplementært til et mRNA-kodon, altsåantall antikodoner, og derfor generelt tRNA-molekyler lik antall mRNA-kodoner;
- mRNA er komplementær til en av DNA-trådene, altsåantall mRNA-nukleotider er lik antall DNA-nukleotider. Antallet trillinger vil selvfølgelig også være det samme.
Når du løser en rekke problemer i denne delen, er det nødvendig å bruke tabellen over den genetiske koden. Reglene for bruk av tabellen er vanligvis angitt i oppgaven, men det er bedre å lære dette på forhånd. For å bestemme aminosyren som er kodet av en bestemt triplett, må du utføre følgende trinn:
- det første nukleotidet til tripletten finnes i den venstre vertikale raden,
- den andre - i den øvre horisontale,
- den tredje er i høyre vertikal rad.
- aminosyren som tilsvarer tripletten ligger i skjæringspunktet mellom de imaginære linjene, kommer fra nukleotider.
1.2. Skriftlig formulering av problemløsning.
Svar:
Et viktig poeng i å løse oppgaver er en forklaring på handlingene som er utført, spesielt hvis oppgaven sier det: «Forklar svaret». Tilstedeværelsen av forklaringer lar verifikatoren konkludere med at studenten forstår emnet, og deres fravær kan føre til tap av en svært viktig poengsum. Oppgave 39 er estimert til tre poeng, som tildeles ved en helt riktig løsning. Derfor, starter oppgaven, først av alt, er det nødvendig å identifisere alle spørsmålene. Antall svar må samsvare.
Kapittel 2. Typer oppgaver i cytologi.
2.1 Oppgaver relatert til å bestemme prosentandelen av nukleotider i DNA
Allerede før oppdagelsen av Watson og Crick, i 1950, slo den australske biokjemikeren Edwin Chargaff fast atat i DNA til enhver organisme er antallet adenylnukleotider lik antall tymidyl, og antall guanylnukleotider er lik antall cytosylnukleotider (A=T, G=C), eller det totale antallet purin nitrogenholdige baser er lik det totale antallet nitrogenholdige pyrimidinbaser (A+G=C+T ).Disse mønstrene kalles "Chargaffs regler".
Faktum er at når en dobbel helix dannes, er den nitrogenholdige basen til adenin i en kjede alltid motsatt av den nitrogenholdige basen til adenin i den andre kjeden, og motsatt av guanin er cytosin, det vil si at DNA-kjedene ser ut til å utfyller hverandre. Og disse sammenkoblede nukleotidene er komplementære til hverandre (fra latin complementum - addisjon).
Hvorfor følges dette prinsippet? For å svare på dette spørsmålet må vi huske den kjemiske naturen til nitrogenholdige heterosykliske baser. Adenin og guanin tilhører puriner, og cytosin og tymin tilhører pyrimidiner, det vil si at bindinger mellom nitrogenholdige baser av samme natur ikke er etablert. I tillegg tilsvarer komplementære baser geometrisk hverandre, dvs. i størrelse og form. Dermed er komplementariteten til nukleotider den kjemiske og geometriske korrespondansen mellom strukturene til molekylene deres til hverandre.
Nitrogenholdige baser inneholder sterkt elektronegative oksygen- og nitrogenatomer, som bærer en delvis negativ ladning, samt hydrogenatomer, som en delvis positiv ladning oppstår på. På grunn av disse partielle ladningene oppstår det hydrogenbindinger mellom nitrogenbasene til de antiparallelle sekvensene til DNA-molekylet.
En oppgave. I et DNA-molekyl er det 23 % adenylnukleotider av det totale antallet nukleotider. Bestem mengden av tymidyl- og cytosylnukleotider.
Svar: T=23%; C=27 %
En oppgave Gitt et DNA-molekyl med en relativ molekylvekt på 69 tusen, hvorav 8625 er adenylnukleotider. Den relative molekylvekten til ett nukleotid er i gjennomsnitt 345. Hvor mange nukleotider er det individuelt i dette DNA? Hva er lengden på molekylet?
Gitt: M(r) DNA - 69000 Antall A - 8625 M(r) nukleotid - 345 Definere: Antall nukleotider i DNA | Løsning : 1. Bestem hvor mange adenylnukleotider som er i et gitt DNA-molekyl: 8625: 345 = 25. 2. Etter Chargaff-regelen er A=G, dvs. i dette DNA-molekylet A=T=25. 3. Bestem hvor mye av den totale molekylvekten til dette DNAet er andelen av guanylnukleotider: 69 000 - (8625x2) = 51 750. 4. Bestem det totale antallet guanyl- og cytosylnukleotider i dette DNA: 51 750:345=150. 5. Bestem innholdet av guanyl- og cytosylnukleotider separat: 150:2 = 75; 6. Bestem lengden på dette DNA-molekylet: (25 + 75) x 0,34 = 34 nm. |
Svar : A=T=25; G=C=75; 34 nm.
2.2. R beregningsoppgaver viet til å bestemme antall aminosyrer i et protein, samt antall nukleotider og tripletter i DNA eller RNA.
- Aminosyrene som kreves for proteinsyntese, leveres til ribosomene via tRNA. Hvert tRNA-molekyl bærer bare én aminosyre.
- Informasjon om primærstrukturen til et proteinmolekyl er kryptert i DNA-molekylet.
- Hver aminosyre er kodet av en sekvens på tre nukleotider. Denne sekvensen kalles en triplett eller kodon.
En oppgave: 30 t-RNA-molekyler deltok i translasjonen. Bestem antall aminosyrer som utgjør det resulterende proteinet, samt antall tripletter og nukleotider i genet som koder for dette proteinet.
Svar : Antall a / c - 30. Antall tripletter - 30. Antall nukleotider - 90.
En oppgave: Ifølge noen forskere er den totale lengden av alle DNA-molekyler i kjernen til én menneskelig kjønnscelle omtrent 102 cm Hvor mange basepar er det i DNA-et til én celle (1 nm = 10–6 mm)?
Svar: 3x109 par.
2.3. Oppgaver for konstruksjon av et i-RNA-molekyl, t-RNA-antikodoner og aminosyresekvenser i en polypeptidkjede. Arbeidmed en tabell over den genetiske koden.
En oppgave: RNA inneholder uracil i stedet for tymin. Proteinbiosyntese involverte t-RNA med antikodoner: UUA, GHC, CHC, AUU, CGU. Bestem nukleotidsekvensen til delen av hver kjede av DNA-molekylet som bærer informasjon om det syntetiserte polypeptidet, og antall nukleotider som inneholder adenin, guanin, tymin, cytosin i et dobbelttrådet DNA-molekyl
Gitt: T-RNA - UUA, GHC, CHC, AUU, CGU | Løsning:
Tall A=T=7, tall G=C=8 |
Definere: Nukleotidsekvens av en del av hver tråd av et DNA-molekyl Antall nukleotider som inneholder adenin, guanin, tymin, cytosin i et DNA-molekyl |
En oppgave: et fragment av en av DNA-kjedene har følgende struktur: AAGGTSTACGTTG. Bygg i-RNA på det og bestem sekvensen av aminosyrer i et fragment av et proteinmolekyl.
Svar: fen-arg-cis-asn.
En oppgave: Ribosomer fra forskjellige celler, hele settet med aminosyrer og de samme molekylene av i-RNA og t-RNA ble plassert i et reagensrør, og skapte alle forutsetninger for proteinsyntese. Hvorfor i et reagensrør vil en type protein syntetiseres på forskjellige ribosomer.
Svar: På samme mRNA syntetiseres det samme proteinet, siden informasjonen er den samme.
- En oppgave: i-RNA-fragmentet har følgende struktur: GAUGAGUATSUUTCAAA. Bestem tRNA-antikodonene og aminosyresekvensen som er kodet i dette fragmentet. Skriv også ned fragmentet av DNA-molekylet som dette mRNA ble syntetisert på.
Svar : Aminosyresekvens - : asp-glu-thir-phen-lys. tRNA-antikodoner - CUA, CUC, AUG, AAG, UUU. DNA-fragment - CTACTCATGAAGTTT
2.4 Oppgaver for å bestemme antall DNA-molekyler og kromosomer i prosessen med mitose og meiose.
Mitose - hovedmetoden for deling av eukaryote celler, der dobling først skjer, og deretter en jevn fordeling mellom dattercellene til arvematerialet.
Mitose er en kontinuerlig prosess der det er fire faser: profase, metafase, anafase og telofase. Før mitose forbereder cellen seg på deling, eller interfase. Perioden med celleforberedelse for mitose og mitose selv utgjør sammenmitotisk syklus.
Interfase består av tre perioder: presyntetisk eller postmitotisk, - G 1 , syntetisk - S, postsyntetisk eller premitotisk - G 2 .
Presyntetisk periode(2 n 2 c , hvor n - antall kromosomer, Med - antall DNA-molekyler) - cellevekst, aktivering av biologiske synteseprosesser, forberedelse til neste periode.
Syntetisk periode(2n4s ) - DNA-replikasjon.
MERK FØLGENDE!
Etter dobling forblir kromosomsettet diploid (2n) fordi søsterkromatidene forblir forbundet ved sentromeren.
Postsyntetisk periode(2n4s ) - forberedelse av cellen for mitose, syntese og akkumulering av proteiner og energi for den kommende divisjonen, en økning i antall organeller, dobling av sentrioler.
Profase (2 n 4 s ) - demontering av kjernemembraner, divergens av sentrioler til forskjellige poler i cellen, dannelse av fisjonsspindeltråder, "forsvinning" av nukleoler, kondensering av to-kromatidkromosomer.
Metafase (2 n 4 s ) - justering av de mest kondenserte to-kromatidkromosomene i ekvatorialplanet til cellen (metafaseplate), festing av spindelfibrene med den ene enden til sentriolene, den andre - til sentromerene til kromosomene.
Anafase (4 n 4 s ) - delingen av to-kromatidkromosomer i kromatider og divergensen av disse søsterkromatidene til motsatte poler i cellen (menskromatider blir uavhengige enkeltkromatidkromosomer).
Telofase (2 n 2 s i hver dattercelle) - dekondensering av kromosomer, dannelse av kjernemembraner rundt hver gruppe kromosomer, desintegrering av fisjonsspindeltrådene, utseendet til nukleolus, delingen av cytoplasma (cytotomi). Cytotomi i dyreceller oppstår på grunn av fisjonsfuren, i planteceller - på grunn av celleplaten.
Meiose - dette er en spesiell måte å dele eukaryote celler på, som et resultat av at overgangen av celler fra en diploid tilstand til en haploid skjer. Meiose består av to påfølgende divisjoner etterfulgt av en enkelt DNA-replikasjon.
Første meiotiske deling (meiose 1)kalt reduksjon, fordi det er under denne delingen at antall kromosomer halveres: fra en diploid celle (2 n 4 s ) danner to haploide (1 n2c).
Mellomfase 1 (i begynnelsen - 2 n 2 c, på slutten - 2 n 4 c ) - syntese og akkumulering av stoffer og energi som er nødvendig for implementering av begge delingene, en økning i cellestørrelse og antall organeller, dobling av sentrioler, DNA-replikasjon, som ender i profeti 1.
Profase 1 (2 n 4 s ) - demontering av kjernemembraner, divergens av sentrioler til ulike poler i cellen, dannelse av fisjonsspindeltråder, "forsvinning" av nukleoler, kondensering av to-kromatide kromosomer, konjugering av homologe kromosomer og kryssing.
Metafase 1 (2 n 4 s ) - justering av bivalente i ekvatorialplanet til cellen, festing av fisjonsspindeltrådene i den ene enden til sentriolene, den andre - til sentromerene til kromosomene.
Anafase 1 (2 n 4 s ) - tilfeldig uavhengigdivergens av to-kromatidkromosomertil motsatte poler av cellen (fra hvert par homologe kromosomer går det ene kromosomet til den ene polen, det andre til den andre), rekombinasjon av kromosomer.
Telofase 1 (1 n 2 s i hver celle) - dannelsen av kjernemembraner rundt grupper av to-kromatidkromosomer, delingen av cytoplasma. I mange planter går en celle fra anafase 1 umiddelbart over til profase 2.
Andre meiotisk deling (meiose 2) kalles likning.
Interfase 2, eller interkinesis (1n 2c ), er en kort pause mellom den første og andre meiotiske delingen der DNA-replikasjon ikke skjer. karakteristisk for dyreceller.
Profase 2 (1 n 2 s ) - demontering av kjernefysiske membraner, divergens av sentrioler til forskjellige poler i cellen, dannelse av fisjonsspindelfilamenter.
Metafase 2 (1 n 2 s ) - justering av to-kromatidkromosomer i ekvatorialplanet til cellen (metafaseplate), festing av spindelfibrene med den ene enden til sentriolene, den andre - til sentromerene til kromosomene; 2 blokker av oogenese hos mennesker.
Anafase 2 (2 n 2 s ) - delingen av to-kromatidkromosomer i kromatider og divergensen av disse søsterkromatidene til motsatte poler av cellen (i dette tilfellet blir kromatidene uavhengige enkeltkromatidkromosomer), rekombinasjon av kromosomer.
Telofase 2 (1 n 1 s i hver celle) - dekondensering av kromosomer, dannelse av kjernemembraner rundt hver gruppe kromosomer, desintegrering av fisjonsspindeltrådene, utseendet til nukleolus, deling av cytoplasma (cytotomy) med dannelse av fire haploide celler som et resultat.

En oppgave: Storfe har 60 kromosomer i sine somatiske celler. Bestem antall kromosomer og DNA-molekyler i eggstokkceller i interfasen før delingen begynner og etter meiose I-deling Forklar hvordan et slikt antall kromosomer og DNA-molekyler dannes.
Svar: Interfase inneholder 60 kromosomer og 120 DNA-molekyler. Etter meiose I - 30 kromosomer og 60 DNA-molekyler.
En oppgave: Kromosomsettet av somatiske hveteceller er 28. Bestem kromosomsettet og antall DNA-molekyler i kjernen (cellen) til eggløsningen før meiose I og meiose II. Forklar resultatene i hvert enkelt tilfelle.
Svar: Før meiose I = 28 kromosomer, 56 DNA-molekyler. Før meiose II = 14 kromosomer, 28 DNA-molekyler
En oppgave: Det er kjent at somatiske celler av kål inneholder 18 kromosomer. Bestem kromosomsettet og antall DNA-molekyler i en av cellene i eggløsningen før starten av meiose, i anafase av meiose I og i anafase av meiose II. Forklar hvilke prosesser som foregår i disse periodene og hvordan de påvirker endringen i antall DNA og kromosomer.
Svar: Før starten av meiose - 18 kromosomer, 36 DNA-molekyler. Anafase I av meiose har 18 kromosomer og 36 DNA-molekyler. I anafase er det 18 kromosomer, 18 DNA-molekyler.
2.5. Oppgaver for å bestemme lengden på en enkelt seksjon av DNA eller antall nukleotider i den.
I følge modellen til den amerikanske biokjemikeren J. Watson og den engelske fysikeren F. Crick, er DNA-molekyler to høyrehendte polynukleotidkjeder rundt en felles akse, eller en dobbel helix. Det er omtrent 10 nukleotidrester per omdreining av helixen. Trådene i denne doble helixen er anti-parallelle, det vil si at de peker i motsatte retninger, slik at 3"-enden av den ene strengen er motsatt 5"-enden av den andre. Størrelsen på DNA-molekyler uttrykkes vanligvis ved antall nukleotider som danner dem. Disse størrelsene varierer fra flere tusen basepar i bakterieplasmider og noen virus til mange hundre tusen basepar i høyere organismer.
Lineær lengde av ett nukleotid i nukleinsyre
L n \u003d 0,34 nm \u003d 3,4 ångstrøm
Gjennomsnittlig molekylvekt av ett nukleotid
Mr n = 345 a.m.u. (Ja)
En oppgave. Konturlengden til et bakteriofag DNA-molekyl er 17x10" 6 m. Etter eksponering for mutagener viste lengden seg å være 13,6x10-6 m. Bestem hvor mange par nitrogenholdige baser som har falt ut som følge av en mutasjon, hvis det er kjent at avstanden mellom tilstøtende nukleotider er 34x10 11 m.
Gitt: Konturlengde på DNA-molekylet - 17x10" 6 m Etter eksponering - 13,6x10-6 m Avstanden mellom tilstøtende nukleotider er -34x10 11 m. | Løsning. 1) Beregn den totale lengden på bakteriofag-DNA-segmentet som har falt ut som følge av eksponering for mutagener. 17 x 10 "6 - 13,6 x 106 \u003d 3,4 x 10 6 (m). 2) Beregn antall nukleotidpar i det droppede fragmentet: 3,4x10-6 / 34x10 "11 \u003d 104 \u003d 10 OOO (par med nukleotider) |
Definer: hvor mange par nitrogenholdige baser som har falt ut som følge av en mutasjon |
Svar: 10 tusen basepar.
Kapittel 4. Eksempler på oppgaver for selvstendig løsning.
- DNA-molekylet inneholder 31 % adenin. Bestem hvor mange (i %) dette molekylet inneholder andre nukleotider.
- Oversettelse involverte 50 t-RNA-molekyler. Bestem antall aminosyrer som utgjør det resulterende proteinet, samt antall tripletter og nukleotider i genet som koder for dette proteinet.
- Et DNA-fragment består av 72 nukleotider. Bestem antall tripletter og nukleotider i mRNA, samt antall aminosyrer som utgjør det resulterende proteinet.
- Et fragment av en av DNA-kjedene har følgende struktur: GGCTCTAGCTTTC. Bygg i-RNA på det og bestem sekvensen av aminosyrer i et fragment av et proteinmolekyl (for dette, bruk tabellen med den genetiske koden).
- i-RNA-fragmentet har følgende struktur: ГЦУАУГУУУУУУУКАЦ. Bestem tRNA-antikodonene og aminosyresekvensen som er kodet i dette fragmentet. Skriv også ned fragmentet av DNA-molekylet som dette mRNA ble syntetisert på (bruk tabellen over den genetiske koden for dette).
- DNA-fragmentet har følgende nukleotidsekvens AGCCGACCTTGCCC. Angi nukleotidsekvensen til t-RNA som syntetiseres på dette fragmentet og aminosyren som dette t-RNA vil bære hvis den tredje tripletten tilsvarer t-RNA-antikodonet. For å løse problemet, bruk tabellen med den genetiske koden.
- I en dyrecelle er det diploide kromosomsettet 20. Bestem antall DNA-molekyler før mitose, etter mitose, etter første og andre deling av meiose.
- 15 glukosemolekyler gikk i dissimilering. Bestem mengden ATP etter glykolyse, etter energistadiet og den totale effekten av dissimilering.
- 6 PVC-molekyler kom inn i Krebs-syklusen. Bestem mengden ATP etter energistadiet, den totale effekten av dissimilering og antall glukosemolekyler som har gått inn i dissimilering.
Svar:
- T=31%, G=C=19% hver.
- 50 aminosyrer, 50 tripletter, 150 nukleotider.
- 24 tripletter, 24 aminosyrer, 24 tRNA-molekyler.
- i-RNA: CCG-AGA-UCG-AAG. Aminosyresekvens: pro-arg-ser-lys.
- DNA-fragment: CGATTACAAGAAATG. tRNA-antikodoner: CGA, UUA, CAA, GAA, AUG. Aminosyresekvens: ala-asn-val-ley-tir.
- t-RNA: UCG-GCU-GAA-CHG. Antikodon GAA, kodon i-RNA - CUU, bærbar aminosyre - leu.
- 2n=20. Genetisk sett:
- før mitose, 40 DNA-molekyler;
- etter mitose, 20 DNA-molekyler;
- etter den første deling av meiose, 20 DNA-molekyler;
- etter andre deling av meiose, 10 DNA-molekyler.
- Siden 2 molekyler av PVC og 2ATP dannes fra ett glukosemolekyl, syntetiseres derfor 30 ATP. Etter energistadiet av dissimilering dannes 36 ATP-molekyler (under nedbrytningen av 1 glukosemolekyl), derfor syntetiseres 540 ATP. Den totale effekten av dissimilering er 540+30=570 ATP.
- 6 PVC-molekyler kom inn i Krebs-syklusen, derfor ble 3 glukosemolekyler spaltet. Mengden ATP etter glykolyse - 6 molekyler, etter energistadiet - 108 molekyler, den totale effekten av dissimilering av 114 ATP-molekyler.
Litteratur:
Kontrolloppgaver i genetikk og retningslinjer for gjennomføring av dem. [komp. L.I. Lushina, S.V. Zalyashchev, A.A. Semenov, O.N. Noskova]. - Samara: SGPU, 2007, 142 s.
http://ege-study.ru
http://licey.net
http://reshuege.ru
http://www.fipi.ru
Vedlegg I genetisk kode (i-RNA)
Første stiftelse | Andre base | Tredje grunn |
|||
hårføner | Ser | Tyr | cis | ||
hårføner | Ser | Tyr | cis | ||
Lei | Ser | ||||
Lei | Ser | Tre | |||
Lei | Pro | gis | Arg | ||
Lei | Pro | gis | Arg | ||
Lei | Pro | Gln | Arg | ||
Lei | Pro | Gln | Arg | ||
ile | Tre | Asn | Ser | ||
ile | Tre | Asn | Ser | ||
ile | Tre | Liz | Arg | ||
Met | Tre | Liz | Arg | ||
Aksel | Ala | Asp | gli | ||
Aksel | Ala | Asp | gli | ||
Aksel | Ala | Glu | gli | ||
Aksel | Ala | Glu | gli |
 Forfatteren av artikkelen er D. A. Solovkov, kandidat for biologiske vitenskaper
Forfatteren av artikkelen er D. A. Solovkov, kandidat for biologiske vitenskaper
Typer oppgaver i cytologi
Oppgavene i cytologi som finnes på eksamen kan deles inn i syv hovedtyper. Den første typen er assosiert med å bestemme prosentandelen av nukleotider i DNA og finnes oftest i del A av eksamen. Den andre gruppen inkluderer beregningsoppgaver viet til å bestemme antall aminosyrer i et protein, samt antall nukleotider og tripletter i DNA eller RNA. Denne typen problemer finnes i både del A og del C.
Oppgaver i cytologi type 3, 4 og 5 er viet arbeid med tabellen over den genetiske koden, og krever også kunnskap om transkripsjons- og oversettelsesprosesser fra søkeren. Slike oppgaver utgjør flertallet av C5-spørsmålene på eksamen.
Oppgaver av type 6 og 7 dukket opp i USE relativt nylig, og de kan søker også støte på i del C. Den sjette typen er basert på kunnskap om endringer i cellens genetiske sett under mitose og meiose, og den syvende type sjekker elevens assimilering av materiale på dissimilering i den eukaryote cellen .
Løsninger på problemer av alle typer er foreslått nedenfor og eksempler for selvstendig arbeid er gitt. Vedlegget inneholder en tabell over den genetiske koden som brukes i løsningen.
Løse problemer av den første typen
Grunnleggende informasjon:
- Det er 4 typer nukleotider i DNA: A (adenin), T (tymin), G (guanin) og C (cytosin).
- I 1953 oppdaget J. Watson og F. Crick at DNA-molekylet er en dobbel helix.
- Kjedene er komplementære til hverandre: i motsetning til adenin i den ene kjeden er det alltid tymin i den andre og omvendt (A-T og T-A); motsatt cytosin - guanin (C-G og G-C).
- I DNA er mengden adenin og guanin lik antall cytosin og tymin, samt A=T og C=G (Chargaffs regel).
Problem: Et DNA-molekyl inneholder adenin. Bestem hvor mange (i) dette molekylet inneholder andre nukleotider.
Løsning: mengden adenin er lik mengden tymin, derfor inneholder dette molekylet tymin. Guanin og cytosin står for  . Fordi deres tall er like, da C=G=.
. Fordi deres tall er like, da C=G=.
Løse problemer av den andre typen
Grunnleggende informasjon:
- Aminosyrene som kreves for proteinsyntese, leveres til ribosomene via tRNA. Hvert tRNA-molekyl bærer bare én aminosyre.
- Informasjon om primærstrukturen til et proteinmolekyl er kryptert i DNA-molekylet.
- Hver aminosyre er kodet av en sekvens på tre nukleotider. Denne sekvensen kalles en triplett eller kodon.
Oppgave: tRNA-molekyler deltok i oversettelsen. Bestem antall aminosyrer som utgjør det resulterende proteinet, samt antall tripletter og nukleotider i genet som koder for dette proteinet.
Løsning: hvis t-RNA var involvert i syntesen, overførte de aminosyrer. Siden én aminosyre er kodet av én triplett, vil det være tripletter eller nukleotider i genet.
Løse problemer av den tredje typen
Grunnleggende informasjon:
- Transkripsjon er prosessen med å syntetisere mRNA fra en DNA-mal.
- Transkripsjon utføres i henhold til regelen om komplementaritet.
- RNA inneholder uracil i stedet for tymin.
Oppgave: et fragment av en av DNA-kjedene har følgende struktur: AAGGCTACGTTTG. Bygg i-RNA på det og bestem sekvensen av aminosyrer i et fragment av et proteinmolekyl.
Løsning: i henhold til komplementaritetsregelen bestemmer vi mRNA-fragmentet og deler det inn i tripletter: UUC-CGA-UHC-AAU. I henhold til tabellen over den genetiske koden bestemmer vi sekvensen av aminosyrer: phen-arg-cis-asn.
Løse problemer av den fjerde typen
Grunnleggende informasjon:
- Et antikodon er en sekvens av tre nukleotider i tRNA som er komplementære til nukleotidene til et mRNA-kodon. tRNA og mRNA inneholder de samme nukleotidene.
- mRNA-molekylet syntetiseres på DNA i henhold til regelen om komplementaritet.
- DNA inneholder tymin i stedet for uracil.
Oppgave: i-RNA-fragment har følgende struktur: GAUGAGUATSUUTCAAA. Bestem tRNA-antikodonene og aminosyresekvensen som er kodet i dette fragmentet. Skriv også ned fragmentet av DNA-molekylet som dette mRNA ble syntetisert på.
Løsning: vi deler mRNA i GAU-GAG-UAC-UUC-AAA-tripletter og bestemmer aminosyresekvensen ved å bruke den genetiske kodetabellen: asp-glu-tir-phen-lys. Dette fragmentet inneholder tripletter, så t-RNA vil delta i syntesen. Deres antikodoner bestemmes i henhold til komplementaritetsregelen: CUA, CUC, AUG, AAG, UUU. I henhold til regelen om komplementaritet bestemmer vi også DNA-fragmentet (ved i-RNA !!!): TSTATSTSATGAAGTTT.
Løse problemer av den femte typen
Grunnleggende informasjon:
- tRNA-molekylet syntetiseres på DNA i henhold til regelen om komplementaritet.
- Husk at RNA inneholder uracil i stedet for tymin.
- Et antikodon er en sekvens av tre nukleotider som er komplementære til nukleotidene til et kodon i mRNA. tRNA og mRNA inneholder de samme nukleotidene.
Oppgave: et DNA-fragment har følgende nukleotidsekvens TTAGCCGATCCG. Angi nukleotidsekvensen til t-RNA som syntetiseres på dette fragmentet og aminosyren som dette t-RNA vil bære hvis den tredje tripletten tilsvarer t-RNA-antikodonet. For å løse problemet, bruk tabellen med den genetiske koden.
Løsning: vi bestemmer sammensetningen av t-RNA-molekylet: AAUCGGCUAGGC og finner den tredje tripletten - dette er CUA. Dette antikodonet er komplementært til i-RNA-tripletten - GAU. Den koder for aminosyren asp, som bæres av dette tRNA.
Løse problemer av den sjette typen
Grunnleggende informasjon:
- De to hovedtypene for celledeling er mitose og meiose.
- Endringer i den genetiske sammensetningen av en celle under mitose og meiose.
Oppgave: i en dyrecelle er det diploide settet med kromosomer lik. Bestem antall DNA-molekyler før mitose, etter mitose, etter første og andre deling av meiose.
Løsning: Etter tilstand, . Genetisk sett:
Løse problemer av den syvende typen
Grunnleggende informasjon:
- Hva er metabolisme, dissimilering og assimilering.
- Dissimilering i aerobe og anaerobe organismer, dens funksjoner.
- Hvor mange stadier i dissimilering, hvor de går, hvilke kjemiske reaksjoner finner sted under hvert trinn.
Oppgave: glukosemolekyler gikk inn i dissimilering. Bestem mengden ATP etter glykolyse, etter energistadiet og den totale effekten av dissimilering.
Løsning: skriv glykolyseligningen: \u003d 2PVC + 4H + 2ATP. Siden PVC- og 2ATP-molekyler dannes fra ett glukosemolekyl, syntetiseres derfor 20 ATP. Etter energistadiet av dissimilering dannes ATP-molekyler (under nedbrytningen av et glukosemolekyl), derfor syntetiseres ATP. Den totale effekten av dissimilering er lik ATP.
Eksempler på oppgaver for selvstendig løsning
- T=, G=C= ved .
- aminosyrer, tripletter, nukleotider.
- triplett, aminosyrer, t-RNA-molekyler.
- i-RNA: CCG-AGA-UCG-AAG. Aminosyresekvens: pro-arg-ser-lys.
- DNA-fragment: CGATTACAAGAAATG. tRNA-antikodoner: CGA, UUA, CAA, GAA, AUG. Aminosyresekvens: ala-asn-val-ley-tir.
- t-RNA: UCG-GCU-GAA-CHG. Antikodonet er GAA, kodonet til i-RNA er CUU, den overførte aminosyren er leu.
- . Genetisk sett:
- Siden PVC- og 2ATP-molekyler dannes fra ett glukosemolekyl, syntetiseres derfor ATP. Etter energistadiet av dissimilering dannes ATP-molekyler (under nedbrytningen av et glukosemolekyl), derfor syntetiseres ATP. Den totale effekten av dissimilering er lik ATP.
- PVC-molekyler kom inn i Krebs-syklusen, derfor brøt glukosemolekyler opp. Mengden ATP etter glykolyse - molekyler, etter energistadiet - molekyler, den totale effekten av dissimilering av ATP-molekyler.
Så denne artikkelen viser hovedtyper av oppgaver i cytologi som en søker kan møte i eksamen i biologi. Vi håper at variantene av oppgavene og deres løsning vil være nyttige for alle i forberedelsene til eksamen. Lykke til!
Celle som et biologisk system
Moderne cellulær teori, dens hovedbestemmelser, rollen i dannelsen av det moderne naturvitenskapelige bildet av verden. Utvikling av kunnskap om cellen. Den cellulære strukturen til organismer er grunnlaget for enheten i den organiske verden, bevis på forholdet mellom levende natur
Moderne cellulær teori, dens hovedbestemmelser, rolle i dannelsen av det moderne naturvitenskapelige bildet av verden
Et av de grunnleggende konseptene i moderne biologi er ideen om at alle levende organismer har en cellulær struktur. Vitenskapen tar for seg studiet av cellens struktur, dens vitale aktivitet og interaksjon med miljøet. cytologi nå ofte referert til som cellebiologi. Cytologi skylder sitt utseende til formuleringen av den cellulære teorien (1838-1839, M. Schleiden, T. Schwann, supplert i 1855 av R. Virchow).
celleteori er en generalisert idé om strukturen og funksjonene til celler som levende enheter, deres reproduksjon og rolle i dannelsen av flercellede organismer.
De viktigste bestemmelsene i celleteorien:
- En celle er en enhet av struktur, livsaktivitet, vekst og utvikling av levende organismer – det er ikke noe liv utenfor cellen.
- En celle er et enkelt system som består av mange elementer som er naturlig forbundet med hverandre, og representerer en viss integrert formasjon.
- Cellene til alle organismer er like i deres kjemiske sammensetning, struktur og funksjoner.
- Nye celler dannes kun som et resultat av deling av moderceller ("celle fra celle").
- Cellene til flercellede organismer danner vev, og organer består av vev. Livet til en organisme som helhet bestemmes av samspillet mellom dens bestanddeler.
- Cellene til flercellede organismer har et komplett sett med gener, men skiller seg fra hverandre ved at ulike grupper av gener jobber for dem, noe som resulterer i det morfologiske og funksjonelle mangfoldet av celler – differensiering.
Takket være opprettelsen av den cellulære teorien ble det klart at cellen er den minste enheten av liv, et elementært levende system, som har alle tegn og egenskaper til levende ting. Formuleringen av den cellulære teorien ble den viktigste forutsetningen for utviklingen av syn på arv og variasjon, siden identifiseringen av deres natur og deres iboende lover uunngåelig antydet universaliteten til strukturen til levende organismer. Å avsløre enheten i den kjemiske sammensetningen og strukturelle planen til cellene fungerte som en drivkraft for utviklingen av ideer om opprinnelsen til levende organismer og deres utvikling. I tillegg har opprinnelsen til flercellede organismer fra en enkelt celle under embryonal utvikling blitt et dogme for moderne embryologi.
Utvikling av kunnskap om cellen
Fram til 1600-tallet visste mennesket overhodet ingenting om mikrostrukturen til gjenstandene rundt seg og oppfattet verden med det blotte øye. Instrumentet for å studere mikroverdenen, mikroskopet, ble oppfunnet omtrent i 1590 av de nederlandske mekanikere G. og Z. Jansen, men dets ufullkommenhet gjorde det umulig å undersøke tilstrekkelig små gjenstander. Bare opprettelsen på grunnlag av det såkalte sammensatte mikroskopet av K. Drebbel (1572-1634) bidro til fremgangen på dette området.
I 1665 forbedret den engelske fysikeren R. Hooke (1635-1703) utformingen av mikroskopet og teknologien for sliping av linser, og for å være sikker på at bildekvaliteten ble bedre, undersøkte han deler av kork, trekull og levende planter under den. På seksjonene fant han de minste porene som lignet en honningkake, og kalte dem celler (fra lat. cellula celle, celle). Det er interessant å merke seg at R. Hooke anså cellemembranen for å være hovedkomponenten i cellen.
I andre halvdel av 1600-tallet dukket opp verkene til de mest fremtredende mikroskopistene M. Malpighi (1628-1694) og N. Gru (1641-1712), som også oppdaget cellestrukturen til mange planter.
For å sikre at det R. Hooke og andre forskere så var sant, utviklet den nederlandske kjøpmannen A. van Leeuwenhoek, som ikke hadde spesialutdanning, uavhengig et mikroskopdesign som var fundamentalt forskjellig fra det eksisterende, og forbedret linseproduksjonen teknologi. Dette tillot ham å oppnå en økning på 275-300 ganger og vurdere slike detaljer i strukturen som var teknisk utilgjengelige for andre forskere. A. van Leeuwenhoek var en uovertruffen observatør: han skisserte og beskrev nøye det han så under et mikroskop, men forsøkte ikke å forklare det. Han oppdaget encellede organismer, inkludert bakterier, fant kjerner, kloroplaster, fortykkelser av cellevegger i planteceller, men oppdagelsene hans kunne evalueres mye senere.
Oppdagelser av komponentene i den indre strukturen til organismer i første halvdel av 1800-tallet fulgte etter hverandre. G. Mol skilte i planteceller levende materie og en vannaktig væske - cellesaft, oppdaget porer. Den engelske botanikeren R. Brown (1773-1858) oppdaget kjernen i orkideceller i 1831, deretter ble den funnet i alle planteceller. Den tsjekkiske vitenskapsmannen J. Purkinje (1787-1869) introduserte begrepet "protoplasma" (1840) for å referere til det halvflytende gelatinøse innholdet i en celle uten kjerne. Den belgiske botanikeren M. Schleiden (1804-1881) kom lenger enn alle sine samtidige, som ved å studere utviklingen og differensieringen av ulike cellulære strukturer av høyere planter, beviste at alle planteorganismer stammer fra én celle. Han vurderte også avrundede kjernelegemer i kjernene til løkskalaceller (1842).
I 1827 oppdaget den russiske embryologen K. Baer eggene til mennesker og andre pattedyr, og tilbakeviste dermed forestillingen om utviklingen av en organisme utelukkende fra mannlige kjønnsceller. I tillegg beviste han dannelsen av en flercellet dyreorganisme fra en enkelt celle - et befruktet egg, samt likheten mellom stadiene av embryonal utvikling av flercellede dyr, noe som antydet enheten av deres opprinnelse. Informasjonen akkumulert ved midten av 1800-tallet krevde generalisering, som ble den cellulære teorien. Biologien skylder sin formulering til den tyske zoologen T. Schwann (1810-1882), som, på grunnlag av sine egne data og konklusjonene til M. Schleiden om utviklingen av planter, foreslo at hvis en kjerne er tilstede i enhver formasjon som er synlig under et mikroskop, så er denne formasjonen celle. Ut fra dette kriteriet formulerte T. Schwann hovedbestemmelsene i celleteorien.
Den tyske legen og patologen R. Virchow (1821-1902) introduserte et annet viktig forslag i denne teorien: celler oppstår kun ved å dele den opprinnelige cellen, det vil si at celler dannes kun fra celler ("celle fra celle").
Siden dannelsen av celleteorien har læren om cellen som en enhet av organismens struktur, funksjon og utvikling blitt kontinuerlig utviklet. På slutten av 1800-tallet, takket være fremskritt innen mikroskopisk teknologi, ble strukturen til cellen avklart, organeller ble beskrevet - deler av cellen som utfører forskjellige funksjoner, metoder for dannelse av nye celler (mitose, meiose) ble studert, og den overordnede betydningen av cellestrukturer i overføringen av arvelige egenskaper ble tydelig. Bruken av de nyeste fysiske og kjemiske forskningsmetoder gjorde det mulig å fordype seg i prosessene for lagring og overføring av arvelig informasjon, samt å studere den fine strukturen til hver av cellestrukturene. Alt dette bidro til separasjonen av vitenskapen om cellen i en uavhengig gren av kunnskap - cytologi.
Den cellulære strukturen til organismer, likheten i strukturen til cellene til alle organismer - grunnlaget for enheten i den organiske verden, bevis på forholdet til levende natur
Alle i dag kjente levende organismer (planter, dyr, sopp og bakterier) har en cellulær struktur. Selv virus som ikke har en cellulær struktur kan bare formere seg i celler. En celle er en elementær strukturell og funksjonell enhet av det levende, som er iboende i alle dens manifestasjoner, spesielt metabolisme og energiomdannelse, homeostase, vekst og utvikling, reproduksjon og irritabilitet. Samtidig er det i cellene arvelig informasjon lagres, behandles og realiseres.
Til tross for alt mangfoldet av celler, er strukturplanen for dem den samme: de inneholder alle arvelig apparatnedsenket i cytoplasma, og den omkringliggende cellen plasmamembran.
Cellen oppsto som et resultat av en lang utvikling av den organiske verden. Foreningen av celler til en flercellet organisme er ikke en enkel summering, siden hver celle, mens den beholder alle egenskapene som er iboende i en levende organisme, samtidig får nye egenskaper på grunn av utførelsen av en viss funksjon av den. På den ene siden kan en flercellet organisme deles inn i dens bestanddeler - celler, men på den annen side, ved å sette dem sammen igjen, er det umulig å gjenopprette funksjonene til en integrert organisme, siden nye egenskaper bare vises i samspillet mellom deler av systemet. Dette manifesterer et av hovedmønstrene som kjennetegner det levende, enheten til det diskrete og det integrerte. Den lille størrelsen og et betydelig antall celler skaper et stort overflateareal i flercellede organismer, noe som er nødvendig for å sikre en rask metabolisme. I tillegg, i tilfelle død av en del av kroppen, kan dens integritet gjenopprettes på grunn av reproduksjon av celler. Utenfor cellen er lagring og overføring av arvelig informasjon, lagring og overføring av energi med dens påfølgende transformasjon til arbeid umulig. Til slutt ga funksjonsdelingen mellom celler i en flercellet organisme store muligheter for organismer til å tilpasse seg omgivelsene og var en forutsetning for komplikasjonen av deres organisasjon.
Dermed tjente etableringen av enheten i planen for strukturen til cellene til alle levende organismer som bevis på enheten til opprinnelsen til alt liv på jorden.
rekke celler. Prokaryote og eukaryote celler. Sammenlignende egenskaper av celler fra planter, dyr, bakterier, sopp Mangfold av celler
I følge den cellulære teorien er en celle den minste strukturelle og funksjonelle enheten av organismer, som har alle egenskapene til en levende ting. I henhold til antall celler deles organismer inn i encellede og flercellede. Celler av encellede organismer eksisterer som uavhengige organismer og utfører alle funksjonene til en levende ting. Alle prokaryoter og en rekke eukaryoter (mange arter av alger, sopp og protozoer) er encellede, som forbløffer med en ekstraordinær variasjon av former og størrelser. Imidlertid er de fleste organismer fortsatt flercellede. Cellene deres er spesialiserte til å utføre visse funksjoner og danne vev og organer, som ikke kan annet enn reflekteres i morfologiske trekk. For eksempel er menneskekroppen dannet av omtrent 10 14 celler, representert av omtrent 200 arter, med et bredt utvalg av former og størrelser.
Formen på cellene kan være rund, sylindrisk, kubisk, prismatisk, skiveformet, spindelformet, stjerneformet, etc. og stjerneformet - celler i nervevevet. En rekke celler har ikke en permanent form i det hele tatt. Disse inkluderer først og fremst blodleukocytter.
Cellestørrelser varierer også betydelig: de fleste celler i en flercellet organisme har størrelser fra 10 til 100 mikron, og de minste - 2-4 mikron. Den nedre grensen skyldes at cellen må ha et minimumssett av stoffer og strukturer for å sikre vital aktivitet, og for store cellestørrelser vil hindre utveksling av stoffer og energi med miljøet, og vil også hindre prosessene for å opprettholde homeostase. Noen celler kan imidlertid sees med det blotte øye. Først av alt inkluderer disse cellene til fruktene til vannmelon og epletrær, samt egg fra fisk og fugler. Selv om en av de lineære dimensjonene til cellen overstiger gjennomsnittet, samsvarer resten med normen. For eksempel kan en nevronutvekst overstige 1 m i lengde, men diameteren vil fortsatt tilsvare gjennomsnittsverdien. Det er ingen direkte sammenheng mellom cellestørrelse og kroppsstørrelse. Så muskelcellene til en elefant og en mus er like store.
Prokaryote og eukaryote celler
Som nevnt ovenfor har celler mange lignende funksjonelle egenskaper og morfologiske egenskaper. Hver av dem består av et cytoplasma nedsenket i den arvelig apparat, og atskilt fra det ytre miljøet plasmamembran, eller plasmalemma, som ikke forstyrrer prosessen med metabolisme og energi. Utenfor membranen kan cellen også ha en cellevegg, bestående av ulike stoffer, som tjener til å beskytte cellen og er et slags ytre skjelett.
Cytoplasmaet er hele innholdet i cellen som fyller rommet mellom plasmamembranen og strukturen som inneholder genetisk informasjon. Den består av hovedstoffet - hyaloplasma- og organeller og inneslutninger nedsenket i den. Organeller- dette er permanente komponenter i cellen som utfører visse funksjoner, og inneslutninger er komponenter som vises og forsvinner i løpet av cellens levetid, og utfører hovedsakelig lagrings- eller utskillelsesfunksjoner. Inneslutninger deles ofte inn i fast og flytende. Faste inneslutninger er hovedsakelig representert av granuler og kan være av en annen karakter, mens vakuoler og fettdråper anses som flytende inneslutninger.

For tiden er det to hovedtyper av celleorganisasjon: prokaryot og eukaryot.
En prokaryot celle har ikke en kjerne; dens genetiske informasjon er ikke atskilt fra cytoplasmaet av membraner.
Området i cytoplasmaet som lagrer genetisk informasjon i en prokaryot celle kalles nukleoid. I cytoplasmaet til prokaryote celler finnes hovedsakelig én type organeller, ribosomer, og organeller omgitt av membraner er helt fraværende. Bakterier er prokaryoter.
En eukaryot celle er en celle der det, i det minste på ett av utviklingsstadiene, er cellekjernen- en spesiell struktur der DNA er lokalisert.
Cytoplasmaet til eukaryote celler kjennetegnes av et betydelig utvalg av membran- og ikke-membranorganeller. Eukaryote organismer inkluderer planter, dyr og sopp. Størrelsen på prokaryote celler er som regel en størrelsesorden mindre enn størrelsen på eukaryote celler. De fleste prokaryoter er encellede organismer, mens eukaryoter er flercellede.
Sammenlignende egenskaper av strukturen til celler av planter, dyr, bakterier og sopp
I tillegg til egenskapene som er karakteristiske for prokaryoter og eukaryoter, har cellene til planter, dyr, sopp og bakterier en rekke andre egenskaper. Så planteceller inneholder spesifikke organeller - kloroplaster, som bestemmer deres evne til fotosyntese, mens i andre organismer finnes ikke disse organellene. Dette betyr selvfølgelig ikke at andre organismer ikke er i stand til fotosyntese, siden det for eksempel i bakterier forekommer på invaginasjoner av plasmalemmaet og individuelle membranvesikler i cytoplasmaet.
Planteceller inneholder vanligvis store vakuoler fylt med cellesaft. I cellene til dyr, sopp og bakterier finnes de også, men de har en helt annen opprinnelse og utfører ulike funksjoner. Hovedreservestoffet som finnes i form av faste inneslutninger er stivelse i planter, glykogen i dyr og sopp, og glykogen eller volutin i bakterier.
Et annet kjennetegn ved disse organismegruppene er organiseringen av overflateapparatet: cellene til dyreorganismer har ikke en cellevegg, plasmamembranen deres er bare dekket med en tynn glykokalyx, mens alle resten har det. Dette er helt forståelig, siden måten dyr mater på er assosiert med fangst av matpartikler i prosessen med fagocytose, og tilstedeværelsen av en cellevegg ville frata dem denne muligheten. Den kjemiske naturen til stoffet som utgjør celleveggen er ikke den samme i forskjellige grupper av levende organismer: hvis det i planter er cellulose, så er det i sopp kitin, og i bakterier er det murein. Sammenlignende egenskaper av strukturen til celler av planter, dyr, sopp og bakterier
| skilt | bakterie | Dyr | Sopp | Planter |
| Fôringsmetode | heterotrofe eller autotrofe | Heterotrofisk | Heterotrofisk | autotrofisk |
| Organisering av arvelig informasjon | prokaryoter | eukaryoter | eukaryoter | eukaryoter |
| DNA lokalisering | Nukleoid, plasmider | kjerne, mitokondrier | kjerne, mitokondrier | Kjerne, mitokondrier, plastider |
| plasmamembran | Det er | Det er | Det er | Det er |
| celleveggen | Mureinovaja | — | Kitinøs | Celluloseholdig |
| Cytoplasma | Det er | Det er | Det er | Det er |
| Organeller | Ribosomer | Membran og ikke-membran, inkludert cellesenteret | Membran og ikke-membran | Membran og ikke-membran, inkludert plastider |
| Bevegelsesorganeller | Flagella og villi | Flagella og flimmerhår | Flagella og flimmerhår | Flagella og flimmerhår |
| Vakuoler | Sjelden | kontraktil, fordøyelseskanal | Noen ganger | Sentralvakuole med cellesaft |
| Inkluderinger | Glykogen, volutin | Glykogen | Glykogen | Stivelse |
Forskjeller i strukturen til cellene til representanter for forskjellige riker av dyreliv er vist i figuren.

Den kjemiske sammensetningen av cellen. Makro- og mikroelementer. Forholdet mellom strukturen og funksjonene til uorganiske og organiske stoffer (proteiner, nukleinsyrer, karbohydrater, lipider, ATP) som utgjør cellen. Kjemikaliers rolle i cellen og menneskekroppen
Den kjemiske sammensetningen av cellen
I sammensetningen av levende organismer er de fleste av de kjemiske elementene i det periodiske systemet for elementer av D. I. Mendeleev, oppdaget til dags dato, funnet. På den ene siden inneholder de ikke et eneste element som ikke ville være i den livløse naturen, og på den annen side er konsentrasjonene deres i kropper av livløs natur og levende organismer betydelig forskjellige.
Disse kjemiske elementene danner uorganiske og organiske stoffer. Til tross for at uorganiske stoffer dominerer i levende organismer, er det organiske stoffer som bestemmer den unike egenskapen til deres kjemiske sammensetning og livets fenomen generelt, siden de syntetiseres hovedsakelig av organismer i prosessen med vital aktivitet og spiller en viktig rolle i reaksjoner.

Vitenskapen tar for seg studiet av den kjemiske sammensetningen av organismer og de kjemiske reaksjonene som finner sted i dem. biokjemi.
Det skal bemerkes at innholdet av kjemikalier i ulike celler og vev kan variere betydelig. For eksempel, mens proteiner dominerer blant organiske forbindelser i dyreceller, dominerer karbohydrater i planteceller.
| Kjemisk element | jordskorpen | Sjøvann | Levende organismer |
| O | 49.2 | 85.8 | 65-75 |
| C | 0.4 | 0.0035 | 15-18 |
| H | 1.0 | 10.67 | 8-10 |
| N | 0.04 | 0.37 | 1.5-3.0 |
| P | 0.1 | 0.003 | 0.20-1.0 |
| S | 0.15 | 0.09 | 0.15-0.2 |
| K | 2.35 | 0.04 | 0.15-0.4 |
| Ca | 3.25 | 0.05 | 0.04-2.0 |
| Cl | 0.2 | 0.06 | 0.05-0.1 |
| mg | 2.35 | 0.14 | 0.02-0.03 |
| Na | 2.4 | 1.14 | 0.02-0.03 |
| Fe | 4.2 | 0.00015 | 0.01-0.015 |
| Zn | < 0.01 | 0.00015 | 0.0003 |
| Cu | < 0.01 | < 0.00001 | 0.0002 |
| Jeg | < 0.01 | 0.000015 | 0.0001 |
| F | 0.1 | 2.07 | 0.0001 |
Makro- og mikroelementer
Omtrent 80 kjemiske grunnstoffer finnes i levende organismer, men bare 27 av disse grunnstoffene har sine funksjoner i cellen og organismen. Resten av elementene er tilstede i spormengder, og ser ut til å bli inntatt gjennom mat, vann og luft. Innholdet av kjemiske elementer i kroppen varierer betydelig. Avhengig av konsentrasjonen er de delt inn i makronæringsstoffer og mikroelementer.
Konsentrasjonen av hver makronæringsstoffer i kroppen overstiger 0,01%, og deres totale innhold er 99%. Makronæringsstoffer inkluderer oksygen, karbon, hydrogen, nitrogen, fosfor, svovel, kalium, kalsium, natrium, klor, magnesium og jern. De fire første av disse grunnstoffene (oksygen, karbon, hydrogen og nitrogen) kalles også organisk, siden de er en del av de viktigste organiske forbindelsene. Fosfor og svovel er også komponenter i en rekke organiske stoffer, som proteiner og nukleinsyrer. Fosfor er avgjørende for dannelsen av bein og tenner.
Uten de gjenværende makronæringsstoffene er normal funksjon av kroppen umulig. Så kalium, natrium og klor er involvert i prosessene for eksitasjon av celler. Kalium er også nødvendig for at mange enzymer skal fungere og holde på vann i cellen. Kalsium finnes i celleveggene til planter, bein, tenner og bløtdyrskjell, og er nødvendig for muskelsammentrekning og intracellulær bevegelse. Magnesium er en komponent av klorofyll - pigmentet som sørger for flyten av fotosyntese. Det tar også del i proteinbiosyntesen. Jern, i tillegg til å være en del av hemoglobin, som bærer oksygen i blodet, er nødvendig for prosessene med respirasjon og fotosyntese, så vel som for funksjonen til mange enzymer.
sporstoffer finnes i kroppen i konsentrasjoner på mindre enn 0,01 %, og deres totale konsentrasjon i cellen når ikke engang 0,1 %. Sporelementer inkluderer sink, kobber, mangan, kobolt, jod, fluor osv. Sink er en del av bukspyttkjertelhormonmolekylet insulin, kobber er nødvendig for fotosyntese og respirasjon. Kobolt er en komponent av vitamin B12, hvis fravær fører til anemi. Jod er nødvendig for syntesen av skjoldbruskhormoner, som sikrer normal metabolisme, og fluor er assosiert med dannelsen av tannemalje.
Både mangel og overskudd eller forstyrrelse av metabolismen av makro- og mikroelementer fører til utvikling av ulike sykdommer. Spesielt forårsaker mangel på kalsium og fosfor rakitt, mangel på nitrogen forårsaker alvorlig proteinmangel, jernmangel forårsaker anemi, og mangel på jod forårsaker brudd på dannelsen av skjoldbruskkjertelhormoner og en reduksjon i stoffskiftet. Å redusere inntaket av fluor med vann og mat i stor grad forårsaker brudd på fornyelsen av tannemaljen og som et resultat en disposisjon for karies. Bly er giftig for nesten alle organismer. Overskuddet forårsaker irreversibel skade på hjernen og sentralnervesystemet, som manifesteres ved tap av syn og hørsel, søvnløshet, nyresvikt, anfall, og kan også føre til lammelser og sykdommer som kreft. Akutt blyforgiftning er ledsaget av plutselige hallusinasjoner og ender i koma og død.
Mangelen på makro- og mikroelementer kan kompenseres ved å øke innholdet i mat og drikkevann, samt ved å ta medisiner. Så jod finnes i sjømat og iodisert salt, kalsium i eggeskall, etc.
Forholdet mellom strukturen og funksjonene til uorganiske og organiske stoffer (proteiner, nukleinsyrer, karbohydrater, lipider, ATP) som utgjør cellen. Kjemikaliers rolle i cellen og menneskekroppen
uorganiske stoffer
De kjemiske elementene i cellen danner forskjellige forbindelser - uorganiske og organiske. De uorganiske stoffene i cellen inkluderer vann, mineralsalter, syrer, etc., og de organiske stoffene inkluderer proteiner, nukleinsyrer, karbohydrater, lipider, ATP, vitaminer, etc.
Vann(H 2 O) - det vanligste uorganiske stoffet i cellen, som har unike fysisk-kjemiske egenskaper. Den har ingen smak, ingen farge, ingen lukt. Tetthet og viskositet for alle stoffer estimeres av vann. Som mange andre stoffer kan vann være i tre aggregeringstilstander: fast (is), flytende og gassformig (damp). Smeltepunktet for vann er $0°$C, kokepunktet er $100°$C, men oppløsning av andre stoffer i vann kan endre disse egenskapene. Varmekapasiteten til vann er også ganske høy - 4200 kJ / mol K, noe som gjør det mulig for det å ta del i prosessene med termoregulering. I et vannmolekyl er hydrogenatomer plassert i en vinkel på $105°$, mens de vanlige elektronparene blir trukket bort av det mer elektronegative oksygenatomet. Dette bestemmer dipolegenskapene til vannmolekyler (en av endene deres er positivt ladet og den andre negativt) og muligheten for dannelse av hydrogenbindinger mellom vannmolekyler. Adhesjonen til vannmolekyler ligger til grunn for fenomenet overflatespenning, kapillaritet og egenskapene til vann som et universelt løsningsmiddel. Som et resultat er alle stoffer delt inn i vannløselige (hydrofile) og uløselige i vann (hydrofob). Takket være disse unike egenskapene er det forhåndsbestemt at vann har blitt grunnlaget for livet på jorden.

Gjennomsnittlig vanninnhold i cellene i kroppen er ikke det samme og kan endre seg med alderen. Så i et halvannen måned gammelt menneskelig embryo når vanninnholdet i cellene 97,5%, hos en åtte måneder gammelt - 83%, hos en nyfødt synker det til 74%, og hos en voksen er det gjennomsnittlig 66%. Imidlertid er kroppsceller forskjellige i vanninnhold. Så beinene inneholder omtrent 20% vann, leveren - 70%, og hjernen - 86%. I det hele tatt kan man si det konsentrasjonen av vann i cellene er direkte proporsjonal med stoffskiftet.
mineralsalter kan være i oppløst eller uoppløst tilstand. Løselige salter dissosiere til ioner - kationer og anioner. De viktigste kationene er kalium- og natriumioner, som letter overføringen av stoffer over membranen og deltar i forekomsten og ledningen av en nerveimpuls; samt kalsiumioner, som deltar i prosessene med sammentrekning av muskelfibre og blodpropp; magnesium, som er en del av klorofyll; jern, som er en del av en rekke proteiner, inkludert hemoglobin. De viktigste anionene er fosfatanionet, som er en del av ATP og nukleinsyrer, og karbonsyreresten som myker opp svingninger i pH i mediet. Ioner av mineralsalter sørger for både penetrering av vann selv inn i cellen og retensjon i den. Hvis konsentrasjonen av salter i miljøet er lavere enn i cellen, trenger vann inn i cellen. Ioner bestemmer også bufferegenskapene til cytoplasmaet, dvs. dens evne til å opprettholde en konstant svakt alkalisk pH i cytoplasmaet, til tross for konstant dannelse av sure og alkaliske produkter i cellen.
Uløselige salter(CaCO 3, Ca 3 (PO 4) 2 osv.) er en del av bein, tenner, skjell og skjell til encellede og flercellede dyr.
I tillegg kan andre uorganiske forbindelser, som syrer og oksider, produseres i organismer. Dermed produserer parietalcellene i menneskets mage saltsyre, som aktiverer fordøyelsesenzymet pepsin, og silisiumoksid impregnerer celleveggene til kjerringrokk og danner kiselalger. De siste årene har også nitrogenoksids (II) rolle i signalisering i celler og kroppen blitt undersøkt.
organisk materiale
Generelle kjennetegn ved de organiske stoffene i cellen
De organiske stoffene i en celle kan representeres av både relativt enkle molekyler og mer komplekse. I tilfeller hvor et komplekst molekyl (makromolekyl) dannes av et betydelig antall repeterende enklere molekyler, kalles det polymer, og strukturelle enheter - monomerer. Avhengig av om enhetene av polymerer gjentas eller ikke, er de klassifisert som regelmessig eller uregelmessig. Polymerer utgjør opptil 90 % av tørrstoffmassen til cellen. De tilhører tre hovedklasser av organiske forbindelser - karbohydrater (polysakkarider), proteiner og nukleinsyrer. Vanlige polymerer er polysakkarider, mens proteiner og nukleinsyrer er uregelmessige. I proteiner og nukleinsyrer er sekvensen av monomerer ekstremt viktig, siden de utfører en informasjonsfunksjon.

Karbohydrater
Karbohydrater- dette er organiske forbindelser, som hovedsakelig inkluderer tre kjemiske elementer - karbon, hydrogen og oksygen, selv om en rekke karbohydrater også inneholder nitrogen eller svovel. Den generelle formelen for karbohydrater er C m (H 2 O) n. De er delt inn i enkle og komplekse karbohydrater.
Enkle karbohydrater (monosakkarider) inneholder et enkelt sukkermolekyl som ikke kan brytes ned til enklere. Dette er krystallinske stoffer, søte på smak og svært løselige i vann. Monosakkarider tar en aktiv del i stoffskiftet i cellen og er en del av komplekse karbohydrater - oligosakkarider og polysakkarider.
Monosakkarider er klassifisert etter antall karbonatomer (C3-C9), for eksempel, pentoser(C 5) og heksoser(Fra 6). Pentoser inkluderer ribose og deoksyribose. Ribose er en del av RNA og ATP. Deoksyribose er en del av DNA. Heksoser (C 6 H 12 O 6) er glukose, fruktose, galaktose, etc. Glukose(druesukker) finnes i alle organismer, inkludert menneskeblod, da det er en energireserve. Det er en del av mange komplekse sukkerarter: sukrose, laktose, maltose, stivelse, cellulose, etc. Fruktose(fruktsukker) finnes i de høyeste konsentrasjonene i frukt, honning, sukkerroerotvekster. Det tar ikke bare en aktiv del i metabolske prosesser, men er også en del av sukrose og noen polysakkarider, for eksempel insulin.
De fleste monosakkarider er i stand til å gi en sølvspeilreaksjon og redusere kobber ved å tilsette Fehlings væske (en blanding av løsninger av kobber(II)sulfat og kalium-natriumtartrat) og koke.

Til oligosakkarider inkluderer karbohydrater dannet av flere monosakkaridrester. De er generelt også svært løselige i vann og er søte på smak. Avhengig av antallet av disse restene skilles disakkarider (to rester), trisakkarider (tre) osv. Disakkarider inkluderer sukrose, laktose, maltose osv. sukrose(bete eller rørsukker) består av rester av glukose og fruktose, det finnes i lagringsorganene til enkelte planter. Spesielt mye sukrose i røttene til sukkerroer og sukkerrør, hvor de oppnås på en industriell måte. Det fungerer som en målestokk for søtheten til karbohydrater. Laktose, eller melkesukker, dannet av rester av glukose og galaktose, funnet i mors- og kumelk. Maltose(maltsukker) består av to glukoserester. Det dannes under nedbrytningen av polysakkarider i plantefrø og i menneskets fordøyelsessystem, og brukes i produksjon av øl.

Polysakkarider er biopolymerer hvis monomerer er mono- eller disakkaridrester. De fleste polysakkarider er uløselige i vann og smaker usøtet. Disse inkluderer stivelse, glykogen, cellulose og kitin. Stivelse– Dette er et hvitt pulveraktig stoff som ikke fuktes av vann, men danner en suspensjon når det brygges med varmt vann – en pasta. Stivelse består faktisk av to polymerer, den mindre forgrenede amylose og den mer forgrenede amylopektin (Figur 2.9). Monomeren av både amylose og amylopektin er glukose. Stivelse er hovedreservestoffet til planter, som akkumuleres i store mengder i frø, frukt, knoller, jordstengler og andre lagringsorganer til planter. En kvalitativ reaksjon på stivelse er en reaksjon med jod, der stivelsen blir blåfiolett.

Glykogen(animalsk stivelse) er et reservepolysakkarid av dyr og sopp, som hos mennesker akkumuleres i de største mengder i muskler og lever. Den er også uløselig i vann og smaker usøtet. Monomeren av glykogen er glukose. Sammenlignet med stivelsesmolekyler er glykogenmolekyler enda mer forgrenet.
Cellulose, eller cellulose, - hovedreferansen polysakkarid av planter. Monomeren av cellulose er glukose. Uforgrenede cellulosemolekyler danner bunter som er en del av celleveggene til planter. Cellulose er grunnlaget for tre, det brukes i konstruksjon, i produksjon av tekstiler, papir, alkohol og mange organiske stoffer. Cellulose er kjemisk inert og løses ikke opp i verken syrer eller alkalier. Det brytes heller ikke ned av enzymene i det menneskelige fordøyelsessystemet, men bakterier i tykktarmen hjelper til med å fordøye det. I tillegg stimulerer fiber sammentrekningen av veggene i mage-tarmkanalen, og bidrar til å forbedre arbeidet.

Kitin er et polysakkarid, hvis monomer er et nitrogenholdig monosakkarid. Det er en del av celleveggene til sopp og leddyrskall. I menneskets fordøyelsessystem er det heller ikke noe enzym for å fordøye kitin, bare noen bakterier har det.
Funksjoner av karbohydrater. Karbohydrater utfører plastiske (konstruksjons-), energi-, lagrings- og støttefunksjoner i cellen. De danner celleveggene til planter og sopp. Energiverdien av nedbrytningen av 1 g karbohydrater er 17,2 kJ. Glukose, fruktose, sukrose, stivelse og glykogen er reservestoffer. Karbohydrater kan også være en del av komplekse lipider og proteiner, og danner glykolipider og glykoproteiner, spesielt i cellemembraner. Ikke mindre viktig er rollen til karbohydrater i intercellulær gjenkjennelse og oppfatning av miljøsignaler, siden de fungerer som reseptorer i sammensetningen av glykoproteiner.
Lipider
Lipider er en kjemisk heterogen gruppe av lavmolekylære stoffer med hydrofobe egenskaper. Disse stoffene er uløselige i vann, danner emulsjoner i det, men er lett løselige i organiske løsemidler. Lipider er fete å ta på, mange av dem etterlater karakteristiske ikke-tørkende spor på papiret. Sammen med proteiner og karbohydrater er de en av hovedkomponentene i cellene. Innholdet av lipider i forskjellige celler er ikke det samme, spesielt mange av dem i frøene og fruktene til noen planter, i leveren, hjertet, blodet.
Avhengig av strukturen til molekylet er lipider delt inn i enkle og komplekse. Til enkel lipider inkluderer nøytrale lipider (fett), voks og steroider. Kompleks lipider inneholder også en annen, ikke-lipid komponent. De viktigste av dem er fosfolipider, glykolipider, etc.
Fett er estere av den treverdige alkoholen glyserol og høyere fettsyrer. De fleste fettsyrer inneholder 14-22 karbonatomer. Blant dem er det både mettede og umettede, det vil si som inneholder dobbeltbindinger. Av de mettede fettsyrene er palmitinsyre og stearinsyre vanligst, og av de umettede fettsyrene oljesyre. Noen umettede fettsyrer syntetiseres ikke i menneskekroppen eller syntetiseres i utilstrekkelige mengder, og er derfor uunnværlige. Glyserolrester danner hydrofile hoder, mens fettsyrerester danner hydrofobe haler.

Fett utfører hovedsakelig en lagringsfunksjon i cellene og fungerer som en energikilde. De er rike på subkutant fettvev, som utfører støtdempende og varmeisolerende funksjoner, og hos vannlevende dyr øker det også oppdriften. Plantefett inneholder for det meste umettede fettsyrer, som et resultat av at de er flytende og kalles oljer. Oljer finnes i frøene til mange planter, som solsikke, soyabønner, raps, etc.
Vokser er estere og blandinger av fettsyrer og fettalkoholer. Hos planter danner de en film på overflaten av bladet, som beskytter mot fordampning, penetrering av patogener, etc. Hos en rekke dyr dekker de kroppen eller tjener til å bygge honningkaker.
Til steroider inkluderer lipider som kolesterol, en essensiell komponent i cellemembraner, samt kjønnshormoner østradiol, testosteron, vitamin D, etc.
Fosfolipider, i tillegg til rester av glyserol og fettsyrer, inneholder en rest av ortofosforsyre. De er en del av cellemembraner og gir sine barriereegenskaper.
Glykolipider er også komponenter av membraner, men innholdet der er lavt. Den ikke-lipide delen av glykolipider er karbohydrater.
Funksjoner av lipider. Lipider utfører plastiske (byggende), energi-, lagrings-, beskyttende, utskillende og regulerende funksjoner i cellen, i tillegg er de vitaminer. Det er en viktig komponent i cellemembraner. Ved spaltning av 1 g lipider frigjøres 38,9 kJ energi. De er avsatt i reservatet i forskjellige organer av planter og dyr. I tillegg beskytter subkutant fettvev indre organer mot hypotermi eller overoppheting, samt sjokk. Den regulerende funksjonen til lipider skyldes det faktum at noen av dem er hormoner. Den fete kroppen til insekter tjener til utskillelse.
Ekorn
Ekorn– Dette er høymolekylære forbindelser, biopolymerer, hvis monomerer er aminosyrer knyttet sammen med peptidbindinger.
aminosyre kalt en organisk forbindelse som har en aminogruppe, en karboksylgruppe og et radikal. Totalt finnes rundt 200 aminosyrer i naturen, som er forskjellige i radikaler og den gjensidige ordningen av funksjonelle grupper, men bare 20 av dem kan være en del av proteiner. Disse aminosyrene kalles proteinogen.

Dessverre kan ikke alle proteinogene aminosyrer syntetiseres i menneskekroppen, så de er delt inn i utskiftbare og uerstattelige. Ikke-essensielle aminosyrer dannes i menneskekroppen i den nødvendige mengden, og uerstattelig- Nei. De må komme fra mat, men kan også delvis syntetiseres av tarmmikroorganismer. Det er 8 fullt essensielle aminosyrer. Disse inkluderer valin, isoleucin, leucin, lysin, metionin, treonin, tryptofan og fenylalanin. Til tross for at absolutt alle proteinogene aminosyrer syntetiseres i planter, er vegetabilske proteiner ufullstendige fordi de ikke inneholder et komplett sett med aminosyrer, dessuten overstiger tilstedeværelsen av protein i de vegetative delene av planter sjelden 1-2% av masse. Derfor er det nødvendig å spise proteiner ikke bare av vegetabilsk, men også av animalsk opprinnelse.
En sekvens av to aminosyrer forbundet med peptidbindinger kalles dipeptid, av tre tripeptid osv. Blant peptidene er det så viktige forbindelser som hormoner (oksytocin, vasopressin), antibiotika osv. En kjede med mer enn tjue aminosyrer kalles polypeptid, og polypeptider som inneholder mer enn 60 aminosyrerester er proteiner.

Nivåer av proteinstrukturell organisering. Proteiner kan ha primære, sekundære, tertiære og kvaternære strukturer.
Primærstrukturen til et protein- dette er lineær aminosyresekvens koblet sammen med en peptidbinding. Den primære strukturen bestemmer til slutt spesifisiteten til proteinet og dets unikhet, siden selv om vi antar at det gjennomsnittlige proteinet inneholder 500 aminosyrerester, så er antallet mulige kombinasjoner 20 500. Derfor er en endring i plasseringen av minst en aminogruppe. syre i primærstrukturen medfører en endring sekundære og høyere strukturer, samt egenskapene til proteinet som helhet.

Strukturelle trekk ved proteinet bestemmer dets romlige pakking - fremveksten av sekundære og tertiære strukturer.
sekundær struktur er det romlige arrangementet av et proteinmolekyl i form spiraler eller folder holdes av hydrogenbindinger mellom oksygen- og hydrogenatomene i peptidgruppene med forskjellige svinger av helixen eller foldene. Mange proteiner inneholder mer eller mindre lange regioner med en sekundær struktur. Dette er for eksempel keratiner av hår og negler, silkefibroin.

Tertiær struktur ekorn ( kule) er også en form for romlig folding av polypeptidkjeden, holdt av hydrofobe, hydrogen, disulfid (S-S) og andre bindinger. Det er karakteristisk for de fleste kroppsproteiner, for eksempel muskelmyoglobin.

Kvartær struktur- den mest komplekse, dannet av flere polypeptidkjeder forbundet hovedsakelig med de samme bindingene som i tertiær (hydrofob, ionisk og hydrogen), samt andre svake interaksjoner. Den kvartære strukturen er karakteristisk for noen få proteiner, som hemoglobin, klorofyll, etc.

Formen på molekylet er fibrillære og kuleformet proteiner. Den første av dem er langstrakte, som for eksempel bindevevskollagen eller hår- og neglekeratiner. Kuleproteiner er i form av en kule (kuler), som muskelmyoglobin.
Enkle og komplekse proteiner. Proteiner kan være enkel og kompleks. Enkle proteiner består kun av aminosyrer, mens kompleks proteiner (lipoproteiner, kromoproteiner, glykoproteiner, nukleoproteiner, etc.) inneholder protein og ikke-proteindeler. Kromoproteiner inneholder en farget ikke-protein del. Disse inkluderer hemoglobin, myoglobin, klorofyll, cytokromer, etc. Således, i sammensetningen av hemoglobin, er hver av de fire polypeptidkjedene til globinproteinet assosiert med en ikke-proteindel - hem, i midten av hvilken det er et jern ion, som gir hemoglobin en rød farge. Ikke-protein del lipoproteiner er et lipid og glykoproteiner- karbohydrater. Både lipoproteiner og glykoproteiner er en del av cellemembraner. Nukleoproteiner er komplekser av proteiner og nukleinsyrer (DNA og RNA). De utfører de viktigste funksjonene i prosessene med lagring og overføring av arvelig informasjon.
Proteinegenskaper. Mange proteiner er svært løselige i vann, men det er noen blant dem som bare løses opp i løsninger av salter, alkalier, syrer eller organiske løsemidler. Strukturen til et proteinmolekyl og dets funksjonelle aktivitet avhenger av miljøforhold. Tapet av et proteinmolekyl av strukturen samtidig som det primære opprettholdes, kalles denaturering.

Denaturering skjer på grunn av endringer i temperatur, pH, atmosfærisk trykk, under påvirkning av syrer, alkalier, salter av tungmetaller, organiske løsningsmidler osv. Den omvendte prosessen med å gjenopprette sekundære og høyere strukturer kalles renaturering, men det er ikke alltid mulig. Den fullstendige nedbrytningen av et proteinmolekyl kalles ødeleggelse.
Proteinfunksjoner. Proteiner utfører en rekke funksjoner i cellen: plastisk (konstruksjon), katalytisk (enzymatisk), energi, signal (reseptor), kontraktil (motorisk), transport, beskyttende, regulatorisk og lagring.
Byggefunksjonen til proteiner er assosiert med deres tilstedeværelse i cellemembraner og strukturelle komponenter i cellen. Energi - på grunn av det faktum at under nedbrytningen av 1 g protein frigjøres 17,2 kJ energi. Membranreseptorproteiner er aktivt involvert i oppfatningen av miljøsignaler og deres overføring gjennom cellen, så vel som i intercellulær gjenkjenning. Uten proteiner er bevegelsen av celler og organismer som helhet umulig, siden de danner grunnlaget for flageller og cilia, og gir også muskelsammentrekning og bevegelse av intracellulære komponenter. I blodet til mennesker og mange dyr frakter proteinet hemoglobin oksygen og deler av karbondioksid, mens andre proteiner transporterer ioner og elektroner. Den beskyttende rollen til proteiner er først og fremst assosiert med immunitet, siden interferonproteinet er i stand til å ødelegge mange virus, og antistoffproteiner hemmer utviklingen av bakterier og andre fremmede stoffer. Det er mange hormoner blant proteiner og peptider, for eksempel bukspyttkjertelhormonet insulin, som regulerer konsentrasjonen av glukose i blodet. I noen organismer kan proteiner lagres i reserve, som i belgfrukter i frø, eller proteinene til et kyllingegg.
Nukleinsyrer
Nukleinsyrer er biopolymerer hvis monomerer er nukleotider. For tiden er to typer nukleinsyrer kjent: ribonukleinsyre (RNA) og deoksyribonukleinsyre (DNA).
Nukleotid dannet av en nitrogenholdig base, en pentosesukkerrest og en fosforsyrerest. Funksjonene til nukleotider bestemmes hovedsakelig av de nitrogenholdige basene som utgjør sammensetningen deres, derfor, selv betinget, er nukleotider utpekt med de første bokstavene i navnene deres. Sammensetningen av nukleotider kan omfatte fem nitrogenholdige baser: adenin (A), guanin (G), tymin (T), uracil (U) og cytosin (C). Pentosene til nukleotidene - ribose og deoksyribose - bestemmer hvilket nukleotid som skal dannes - ribonukleotid eller deoksyribonukleotid. Ribonukleotider er RNA-monomerer, de kan fungere som signalmolekyler (cAMP) og være en del av høyenergiforbindelser, som ATP, og koenzymer, som NADP, NAD, FAD osv., og deoksyribonukleotider er en del av DNA.
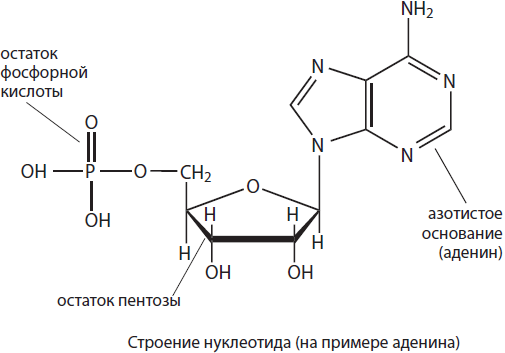
Deoksyribonukleinsyre (DNA)- dobbelttrådet biopolymer, hvis monomerer er deoksyribonukleotider. Sammensetningen av deoksyribonukleotider inkluderer bare fire nitrogenholdige baser av fem mulige - adenin (A), tymin (T), guanin (G) eller cytosin (C), samt deoksyribose og fosforsyrerester. Nukleotider i DNA-kjeden er sammenkoblet gjennom ortofosforsyrerester, og danner en fosfodiesterbinding. Når et dobbelttrådet molekyl dannes, blir nitrogenbasene rettet innover molekylet. Sammenkoblingen av DNA-kjeder skjer imidlertid ikke tilfeldig - de nitrogenholdige basene til forskjellige kjeder er sammenkoblet med hydrogenbindinger i henhold til komplementaritetsprinsippet: adenin er koblet til tymin med to hydrogenbindinger (A \u003d T), og guanin og cytosin med tre (G $ ≡ $ C).

For henne ble satt Chargaff regler:
- Antallet DNA-nukleotider som inneholder adenin er lik antallet nukleotider som inneholder tymin (A=T).
- Antall DNA-nukleotider som inneholder guanin er lik antallet nukleotider som inneholder cytosin (G$≡$C).
- Summen av deoksyribonukleotider som inneholder adenin og guanin er lik summen av deoksyribonukleotider som inneholder tymin og cytosin (A+G = T+C).
- Forholdet mellom summen av deoksyribonukleotider som inneholder adenin og tymin og summen av deoksyribonukleotider som inneholder guanin og cytosin avhenger av typen organisme.
Strukturen til DNA ble dechiffrert av F. Crick og D. Watson (Nobelprisen i fysiologi eller medisin, 1962). I følge deres modell er DNA-molekylet en høyrehendt dobbelhelix. Avstanden mellom nukleotidene i DNA-kjeden er 0,34 nm.

Den viktigste egenskapen til DNA er evnen til å replikere (selvdobling). Hovedfunksjonen til DNA er lagring og overføring av arvelig informasjon, som er skrevet i form av nukleotidsekvenser. Stabiliteten til DNA-molekylet opprettholdes av kraftige reparasjons- (gjenopprettingssystemer), men selv de er ikke i stand til å eliminere uønskede effekter fullstendig, noe som til slutt fører til mutasjoner. DNA fra eukaryote celler er konsentrert i kjernen, mitokondriene og plastidene, mens prokaryote celler befinner seg direkte i cytoplasmaet. Kjernefysisk DNA er grunnlaget for kromosomer, det er representert av åpne molekyler. DNA fra mitokondrier, plastider og prokaryoter har en sirkulær form.
Ribonukleinsyre (RNA)- en biopolymer hvis monomerer er ribonukleotider. De inneholder også fire nitrogenholdige baser - adenin (A), uracil (U), guanin (G) eller cytosin (C), og skiller seg dermed fra DNA i en av basene (i stedet for tymin inneholder RNA uracil). Pentosesukkerresten i ribonukleotider er representert ved ribose. RNA er stort sett enkelttrådede molekyler, med unntak av noen virale. Det er tre hovedtyper av RNA: informasjons- eller mal (mRNA, mRNA), ribosomalt (rRNA) og transport (tRNA). Alle dannes i prosessen transkripsjoner- omskriving fra DNA-molekyler.
og RNA utgjør den minste fraksjon av RNA i en celle (2-4%), noe som oppveies av deres mangfold, siden en celle kan inneholde tusenvis av forskjellige mRNA. Dette er enkelttrådede molekyler som er maler for syntesen av polypeptidkjeder. Informasjon om strukturen til proteinet er registrert i dem i form av sekvenser av nukleotider, og hver aminosyre koder for en triplett av nukleotider - kodon.
R RNA er den mest tallrike typen RNA i cellen (opptil 80%). Deres molekylvekt er gjennomsnittlig 3000-5000; dannes i nukleolene og er en del av celleorganellene - ribosomer. rRNA ser også ut til å spille en rolle i proteinsyntese.
t RNA er det minste av RNA-molekylene, siden det bare inneholder 73-85 nukleotider. Deres andel av den totale mengden celle-RNA er omtrent 16 %. Funksjonen til tRNA er transport av aminosyrer til stedet for proteinsyntese (på ribosomer). Formen på tRNA-molekylet ligner et kløverblad. I den ene enden av molekylet er det et sted for å feste en aminosyre, og i en av løkkene er det en triplett av nukleotider som er komplementær til mRNA-kodonet og bestemmer hvilken aminosyre tRNA skal bære - antikodon.
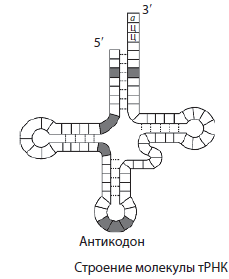
Alle typer RNA deltar aktivt i implementeringen av arvelig informasjon, som skrives om fra DNA til mRNA, og på sistnevnte utføres proteinsyntese. tRNA i prosessen med proteinsyntese leverer aminosyrer til ribosomer, og rRNA er en del av ribosomene direkte.
Adenosintrifosforsyre (ATP) er et nukleotid som inneholder, i tillegg til den nitrogenholdige basen av adenin og en riboserest, tre fosforsyrerester. Bindingene mellom de to siste fosforrestene er makroerge (42 kJ/mol energi frigjøres ved spaltning), mens standard kjemisk binding ved spaltning gir 12 kJ/mol. Hvis energi er nødvendig, deles den makroerge bindingen til ATP, adenosindifosforsyre (ADP), en fosforrest dannes, og energi frigjøres:
ATP + H2O $→$ ADP + H3PO4 + 42 kJ.
ADP kan også brytes ned for å danne AMP (adenosinmonofosforsyre) og en fosforsyrerest:
ADP + H 2 O $ → $ AMP + H 3 PO 4 + 42 kJ.
I prosessen med energimetabolisme (under respirasjon, gjæring), så vel som i prosessen med fotosyntese, fester ADP en fosforrest og blir til ATP. ATP-gjenopprettingsreaksjonen kalles fosforylering. ATP er en universell energikilde for alle livsprosesser til levende organismer.
Studiet av den kjemiske sammensetningen av cellene til alle levende organismer har vist at de inneholder de samme kjemiske elementene, kjemikalier som utfører de samme funksjonene. Dessuten vil et stykke DNA overført fra en organisme til en annen fungere i den, og et protein syntetisert av bakterier eller sopp vil fungere som et hormon eller enzym i menneskekroppen. Dette er et av bevisene på enheten i opprinnelsen til den organiske verden.

Cellestruktur. Forholdet mellom strukturen og funksjonene til delene og organellene i cellen er grunnlaget for dens integritet
Cellestruktur
Strukturen til prokaryote og eukaryote celler
De viktigste strukturelle komponentene til celler er plasmamembranen, cytoplasma og arvelig apparat. Avhengig av egenskapene til organisasjonen, skilles to hovedtyper av celler: prokaryote og eukaryote. Hovedforskjellen mellom prokaryote og eukaryote celler er organiseringen av deres arvelige apparat: i prokaryoter er det lokalisert direkte i cytoplasmaet (dette området av cytoplasmaet kalles nukleoid) og er ikke separert fra den av membranstrukturer, mens i eukaryoter er mesteparten av DNA konsentrert i kjernen, omgitt av en dobbel membran. I tillegg er den genetiske informasjonen til prokaryote celler, lokalisert i nukleoiden, registrert i det sirkulære DNA-molekylet, mens i eukaryoter er DNA-molekylene ikke lukket.
I motsetning til eukaryoter inneholder cytoplasmaet til prokaryote celler også en liten mengde organeller, mens eukaryote celler er preget av en betydelig variasjon av disse strukturene.
Strukturen og funksjonene til biologiske membraner
Strukturen til biomembranen. De cellegrensende membranene og membranorganellene til eukaryote celler deler en felles kjemisk sammensetning og struktur. De inkluderer lipider, proteiner og karbohydrater. Membranlipider er hovedsakelig representert av fosfolipider og kolesterol. De fleste membranproteiner er komplekse proteiner som glykoproteiner. Karbohydrater oppstår ikke av seg selv i membranen, de er assosiert med proteiner og lipider. Tykkelsen på membranene er 7-10 nm.
I henhold til den for tiden aksepterte flytende mosaikkmodellen av membranstruktur, danner lipider et dobbeltlag, eller lipid dobbeltlag, der de hydrofile "hodene" til lipidmolekyler er vendt utover, og de hydrofobe "halene" er skjult inne i membranen. Disse "halene", på grunn av deres hydrofobisitet, sikrer separasjon av de vandige fasene i det indre miljøet til cellen og dens miljø. Proteiner er assosiert med lipider gjennom ulike typer interaksjoner. Noen av proteinene er lokalisert på overflaten av membranen. Slike proteiner kalles perifert, eller overfladisk. Andre proteiner er helt eller delvis nedsenket i membranen - disse er integrert, eller nedsenkede proteiner. Membranproteiner utfører strukturelle, transport-, katalytiske, reseptor- og andre funksjoner.
Membraner er ikke som krystaller, komponentene deres er konstant i bevegelse, som et resultat av at det oppstår hull mellom lipidmolekyler - porer som forskjellige stoffer kan komme inn i eller forlate cellen gjennom.
Biologiske membraner er forskjellige i deres plassering i cellen, deres kjemiske sammensetning og deres funksjoner. Hovedtypene av membraner er plasma og interne. plasmamembran inneholder ca. 45 % lipider (inkludert glykolipider), 50 % proteiner og 5 % karbohydrater. Kjeder av karbohydrater som utgjør komplekse proteiner-glykoproteiner og komplekse lipider-glykolipider stikker ut over overflaten av membranen. Plasmalemmale glykoproteiner er ekstremt spesifikke. Så for eksempel gjennom dem er det en gjensidig anerkjennelse av celler, inkludert sæd og egg.
På overflaten av dyreceller danner karbohydratkjeder et tynt overflatelag - glykokalyx. Det er funnet i nesten alle dyreceller, men alvorlighetsgraden er ikke den samme (10-50 mikron). Glykokalyxen gir en direkte forbindelse mellom cellen og det ytre miljøet, ekstracellulær fordøyelse skjer i den; reseptorer er lokalisert i glykokalyxen. Cellene til bakterier, planter og sopp er i tillegg til plasmalemmaet også omgitt av cellemembraner.
Innvendige membraner eukaryote celler avgrenser forskjellige deler av cellen, og danner en slags "rom" - avdelinger, som bidrar til separasjon av ulike prosesser av metabolisme og energi. De kan variere i kjemisk sammensetning og funksjoner, men de beholder den generelle planen for strukturen.
Membranfunksjoner:
- Begrensende. Det består i det faktum at de skiller det indre rommet i cellen fra det ytre miljøet. Membranen er semipermeabel, det vil si at bare de stoffene som er nødvendige for cellen kan fritt overvinne den, mens det er mekanismer for å transportere de nødvendige stoffene.
- Reseptor. Det er først og fremst assosiert med oppfatningen av miljøsignaler og overføringen av denne informasjonen inn i cellen. Spesielle reseptorproteiner er ansvarlige for denne funksjonen. Membranproteiner er også ansvarlige for cellulær gjenkjennelse i henhold til "venn eller fiende"-prinsippet, samt for dannelsen av intercellulære forbindelser, hvorav de mest studerte er synapsene til nerveceller.
- katalytisk. Tallrike enzymkomplekser er lokalisert på membranene, som et resultat av hvilke intensive syntetiske prosesser finner sted på dem.
- Energitransformasjon. Assosiert med dannelsen av energi, dens lagring i form av ATP og utgifter.
- Oppdeling. Membranene avgrenser også rommet inne i cellen, og skiller derved de innledende stoffene i reaksjonen og enzymene som kan utføre de tilsvarende reaksjonene.
- Dannelse av intercellulære kontakter. Til tross for at membrantykkelsen er så liten at den ikke kan skilles med det blotte øye, fungerer den på den ene siden som en ganske pålitelig barriere for ioner og molekyler, spesielt vannløselige, og på den andre siden. sikrer deres overføring inn i cellen og ut.
- Transportere.

membrantransport. På grunn av det faktum at celler, som elementære biologiske systemer, er åpne systemer, for å sikre metabolisme og energi, opprettholde homeostase, vekst, irritabilitet og andre prosesser, kreves overføring av stoffer gjennom membranen - membrantransport. For tiden er transporten av stoffer over cellemembranen delt inn i aktiv, passiv, endo- og eksocytose.
Passiv transport er en type transport som skjer uten forbruk av energi fra en høyere konsentrasjon til en lavere. Lipidløselige små ikke-polare molekyler (O 2, CO 2) trenger lett inn i cellen ved å enkel diffusjon. Uløselige i lipider, inkludert ladede små partikler, plukkes opp av bærerproteiner eller passerer gjennom spesielle kanaler (glukose, aminosyrer, K+, PO 4 3-). Denne typen passiv transport kalles forenklet diffusjon. Vann kommer inn i cellen gjennom porene i lipidfasen, samt gjennom spesielle kanaler foret med proteiner. Transport av vann over en membran kalles osmose.

Osmose er ekstremt viktig i cellens liv, fordi hvis den plasseres i en løsning med høyere konsentrasjon av salter enn i celleløsningen, vil vann begynne å forlate cellen, og volumet av levende innhold vil begynne å avta . I dyreceller krymper cellen som helhet, og i planteceller henger cytoplasmaet etter celleveggen, som kalles plasmolyse. Når en celle plasseres i en løsning mindre konsentrert enn cytoplasmaet, transporteres vann i motsatt retning – inn i cellen. Det er imidlertid grenser for utvidbarheten til den cytoplasmatiske membranen, og dyrecellen sprekker til slutt, mens i plantecellen tillates dette ikke av en sterk cellevegg. Fenomenet med å fylle hele det indre rommet i cellen med cellulært innhold kalles deplasmolyse. Intracellulær saltkonsentrasjon bør tas i betraktning ved tilberedning av legemidler, spesielt for intravenøs administrering, da dette kan føre til skade på blodceller (til dette brukes fysiologisk saltvann med en konsentrasjon på 0,9 % natriumklorid). Dette er ikke mindre viktig i dyrking av celler og vev, så vel som organer fra dyr og planter.

aktiv transport fortsetter med forbruket av ATP-energi fra en lavere konsentrasjon av et stoff til en høyere. Det utføres ved hjelp av spesielle protein-pumper. Proteiner pumper ioner K +, Na +, Ca 2+ og andre gjennom membranen, noe som bidrar til transport av de viktigste organiske stoffene, samt fremveksten av nerveimpulser mv.
Endocytose- dette er en aktiv prosess for absorpsjon av stoffer av cellen, der membranen danner invaginasjoner, og deretter danner membranvesikler - fagosomer, som inneholder absorberte gjenstander. Det primære lysosomet smelter deretter sammen med fagosomet for å dannes sekundært lysosom, eller fagolysosom, eller fordøyelsesvakuol. Innholdet i vesikkelen spaltes av lysosomenzymer, og spaltningsproduktene absorberes og assimileres av cellen. Ufordøyde rester fjernes fra cellen ved eksocytose. Det er to hovedtyper av endocytose: fagocytose og pinocytose.
Fagocytose er prosessen med fangst av celleoverflaten og absorpsjon av faste partikler av cellen, og pinocytose- væsker. Fagocytose forekommer hovedsakelig i dyreceller (encellede dyr, humane leukocytter), det gir deres ernæring, og ofte beskyttelse av kroppen. Ved hjelp av pinocytose skjer absorpsjon av proteiner, antigen-antistoffkomplekser i prosessen med immunreaksjoner etc. Men mange virus kommer også inn i cellen ved hjelp av pinocytose eller fagocytose. I cellene til planter og sopp er fagocytose praktisk talt umulig, siden de er omgitt av sterke cellemembraner.

Eksocytose er den omvendte prosessen med endocytose. Dermed frigjøres ufordøyde matrester fra fordøyelsesvakuolene, stoffene som er nødvendige for cellens liv og organismen som helhet fjernes. For eksempel skjer overføring av nerveimpulser på grunn av frigjøring av kjemiske budbringere fra nevronet som sender impulsen - formidlere, og i planteceller frigjøres hjelpekarbohydrater i cellemembranen på denne måten.
Cellevegger til planteceller, sopp og bakterier. Utenfor membranen kan cellen skille ut et sterkt rammeverk - cellemembran, eller celleveggen.
Hos planter er celleveggen bygd opp av cellulose pakket i bunter på 50-100 molekyler. Hullene mellom dem er fylt med vann og andre karbohydrater. Plantecellemembranen er gjennomboret av tubuli - plasmodesmata som membranene i det endoplasmatiske retikulum passerer gjennom. Plasmodesmata transporterer stoffer mellom cellene. Imidlertid kan transport av stoffer, som vann, også skje langs selve celleveggene. Over tid akkumuleres ulike stoffer, inkludert tanniner eller fettlignende stoffer, i cellemembranen til planter, noe som fører til lignifisering eller korking av selve celleveggen, forskyvning av vann og død av celleinnhold. Mellom celleveggene til naboplanteceller er det gelélignende puter - midtplater som fester dem sammen og sementerer plantekroppen som helhet. De blir ødelagt bare i prosessen med fruktmodning og når bladene faller.

Celleveggene til soppceller dannes kitin- et karbohydrat som inneholder nitrogen. De er sterke nok og er det ytre skjelettet til cellen, men likevel, som hos planter, forhindrer de fagocytose.
Hos bakterier inneholder celleveggen et karbohydrat med fragmenter av peptider - murein, men innholdet varierer betydelig i ulike grupper av bakterier. På toppen av celleveggen kan også andre polysakkarider frigjøres, og danner en slimete kapsel som beskytter bakterier mot ytre påvirkninger.
Skallet bestemmer cellens form, fungerer som en mekanisk støtte, utfører en beskyttende funksjon, gir cellens osmotiske egenskaper, begrenser strekkingen av det levende innholdet og forhindrer brudd på cellen, som øker på grunn av tilstrømningen av vann. I tillegg overvinner vann og stoffer som er oppløst i det celleveggen før de kommer inn i cytoplasmaet eller omvendt når de forlater det, mens vann transporteres langs celleveggene raskere enn gjennom cytoplasmaet.
Cytoplasma
Cytoplasma er det indre av cellen. Alle organeller i cellen, kjernen og ulike avfallsprodukter er nedsenket i den.
Cytoplasmaet forbinder alle deler av cellen med hverandre, mange metabolske reaksjoner finner sted i den. Cytoplasmaet er skilt fra miljøet og delt inn i rom av membraner, det vil si at celler har en membranstruktur. Det kan være i to tilstander - sol og gel. Sol- dette er en halvflytende, gelélignende tilstand i cytoplasmaet, der vitale prosesser foregår mest intensivt, og gel- en tettere, gelatinøs tilstand som hindrer flyten av kjemiske reaksjoner og transport av stoffer.
Den flytende delen av cytoplasmaet uten organeller kalles hyaloplasma. Hyaloplasma, eller cytosol, er en kolloid løsning der det er en slags suspensjon av ganske store partikler, som proteiner, omgitt av dipoler av vannmolekyler. Sedimenteringen av denne suspensjonen skjer ikke på grunn av at de har samme ladning og frastøter hverandre.
Organeller
Organeller– Dette er permanente komponenter i cellen som utfører visse funksjoner.
Avhengig av de strukturelle egenskapene er de delt inn i membran og ikke-membran. Membran organeller blir på sin side referert til som enkeltmembran (endoplasmatisk retikulum, Golgi-kompleks og lysosomer) eller dobbeltmembran (mitokondrier, plastider og kjerne). Ikke-membran organeller er ribosomer, mikrotubuli, mikrofilamenter og cellesenteret. Av de listede organellene er det bare ribosomer som er iboende i prokaryoter.
Strukturen og funksjonene til kjernen. Cellekjernen- en stor to-membran organell som ligger i midten av cellen eller på dens periferi. Størrelsen på kjernen kan variere innen 3-35 mikron. Formen på kjernen er oftere sfærisk eller ellipsoid, men det er også stavformede, spindelformede, bønneformede, flikete og til og med segmenterte kjerner. Noen forskere mener at formen på kjernen tilsvarer formen på selve cellen.
De fleste celler har én kjerne, men for eksempel i lever- og hjerteceller kan det være to, og i en rekke nevroner – opptil 15. Skjelettmuskelfibre inneholder vanligvis mange kjerner, men de er ikke celler i den fulle forstand av ordet, siden de er dannet som et resultat av fusjon av flere celler.
Kjernen er omgitt kjernefysisk konvolutt, og dens indre plass er fylt atomjuice, eller nukleoplasma (karyoplasma) som er nedsenket i kromatin og nukleolus. Kjernen utfører så viktige funksjoner som lagring og overføring av arvelig informasjon, samt kontroll av cellens vitale aktivitet.

Kjernens rolle i overføringen av arvelig informasjon er overbevisende bevist i eksperimenter med grønnalgen acetabularia. I en enkelt gigantisk celle, som når en lengde på 5 cm, skilles en hatt, et ben og en rhizoid. Dessuten inneholder den bare en kjerne som ligger i rhizoiden. På 1930-tallet transplanterte I. Hemmerling kjernen til en art av acetabularia med en grønn farge til en rhizoide av en annen art, med en brun farge, der kjernen ble fjernet. Etter en tid fikk planten med den transplanterte kjernen en ny hette, som algedonoren til kjernen. Samtidig døde hetten eller stilken fra rhizoiden, som ikke inneholdt en kjerne, etter en tid.

kjernefysisk konvolutt Den er dannet av to membraner - ytre og indre, mellom hvilke det er et mellomrom. Intermembranrommet kommuniserer med hulrommet til det grove endoplasmatiske retikulumet, og den ytre membranen til kjernen kan bære ribosomer. Den kjernefysiske konvolutten er gjennomsyret av mange porer, kantet med spesielle proteiner. Stoffer transporteres gjennom porene: de nødvendige proteinene (inkludert enzymer), ioner, nukleotider og andre stoffer kommer inn i kjernen, og RNA-molekyler, avfallsproteiner, underenheter av ribosomer forlater den. Dermed er funksjonene til kjernekappen separasjon av innholdet i kjernen fra cytoplasma, samt regulering av metabolismen mellom kjernen og cytoplasma.
Nukleoplasma kalt innholdet i kjernen, der kromatinet og nukleolen er nedsenket. Det er en kolloidal løsning, som kjemisk minner om cytoplasmaet. Enzymer i nukleoplasmaet katalyserer utvekslingen av aminosyrer, nukleotider, proteiner osv. Nukleoplasmaet er forbundet med hyaloplasmaet gjennom kjerneporer. Funksjonene til nukleoplasma, som hyaloplasma, er å sikre sammenkobling av alle strukturelle komponenter i kjernen og implementering av en rekke enzymatiske reaksjoner.
kromatin kalt et sett med tynne tråder og granuler nedsenket i nukleoplasmaet. Det kan bare oppdages ved farging, siden brytningsindeksene for kromatin og nukleoplasma er omtrent de samme. Den filamentøse komponenten av kromatin kalles eukromatin og granulert heterokromatin. Eukromatin er svakt komprimert, siden arvelig informasjon leses fra det, mens mer spiralisert heterokromatin er genetisk inaktivt.
Kromatin er en strukturell modifikasjon av kromosomer i en ikke-delende kjerne. Dermed er kromosomer konstant tilstede i kjernen; bare tilstanden deres endres avhengig av funksjonen som kjernen utfører for øyeblikket.
Sammensetningen av kromatin inkluderer hovedsakelig nukleoproteiner (deoksyribonukleoproteiner og ribonukleoproteiner), samt enzymer, hvorav de viktigste er assosiert med syntesen av nukleinsyrer, og noen andre stoffer.
Funksjonene til kromatin består for det første i syntesen av nukleinsyrer spesifikke for en gitt organisme, som styrer syntesen av spesifikke proteiner, og for det andre i overføring av arvelige egenskaper fra modercellen til datterceller, for hvilke kromatintråder er pakket inn i kromosomer under deling.
nukleolus- et sfærisk legeme, godt synlig under et mikroskop med en diameter på 1-3 mikron. Det dannes i kromatinregioner som koder for informasjon om strukturen til rRNA og ribosomproteiner. Nukleolen i kjernen er ofte én, men i de cellene hvor det foregår intensive vitale prosesser, kan det være to eller flere nukleoler. Funksjonene til nukleolene er syntese av rRNA og sammenstilling av ribosomunderenheter ved å kombinere rRNA med proteiner som kommer fra cytoplasmaet.
Mitokondrier- to-membranorganeller av en rund, oval eller stavformet form, selv om spiralformede også finnes (i sædceller). Mitokondrier er opptil 1 µm i diameter og opptil 7 µm i lengde. Rommet inne i mitokondriene er fylt med matrise. Matrise Det er hovedstoffet i mitokondrier. Et sirkulært DNA-molekyl og ribosomer er nedsenket i det. Den ytre membranen til mitokondrier er glatt og ugjennomtrengelig for mange stoffer. Den indre membranen har utvekster - cristae, som øker overflaten av membraner for at kjemiske reaksjoner skal oppstå. På overflaten av membranen er det mange proteinkomplekser som utgjør den såkalte respirasjonskjeden, samt soppformede enzymer av ATP-syntetase. I mitokondrier finner det aerobe respirasjonsstadiet sted, hvor ATP syntetiseres.

plastider- store to-membran organeller, karakteristiske bare for planteceller. Det indre rommet av plastider er fylt stroma, eller matrise. I stroma er det et mer eller mindre utviklet system av membranvesikler - thylakoider, som er samlet i hauger - korn, samt sitt eget sirkulære DNA-molekyl og ribosomer. Det er fire hovedtyper av plastider: kloroplaster, kromoplaster, leukoplaster og proplastider.
Kloroplaster– Dette er grønne plastider med en diameter på 3-10 mikron, godt synlige i mikroskop. De finnes bare i de grønne delene av planter - blader, unge stilker, blomster og frukt. Kloroplaster er for det meste ovale eller ellipsoide i form, men kan også være skålformede, spiralformede og til og med flikete. Antallet kloroplaster i en celle er i gjennomsnitt fra 10 til 100 stykker. Men, for eksempel, i noen alger kan det være en, ha en betydelig størrelse og kompleks form - da kalles det kromatofor. I andre tilfeller kan antallet kloroplaster komme opp i flere hundre, mens størrelsen er liten. Fargen på kloroplaster skyldes hovedpigmentet i fotosyntesen - klorofyll, selv om de inneholder flere pigmenter - karotenoider. Karotenoider blir merkbare først om høsten, når klorofyllet i aldrende blader blir ødelagt. Hovedfunksjonen til kloroplaster er fotosyntese. Lysreaksjoner av fotosyntese forekommer på thylakoidmembraner, hvor klorofyllmolekyler er festet, og mørkereaksjoner forekommer i stroma, som inneholder mange enzymer.
Kromoplaster er gule, oransje og røde plastider som inneholder karotenoidpigmenter. Formen på kromoplaster kan også variere betydelig: de er rørformede, sfæriske, krystallinske osv. Kromoplaster gir farge til blomster og frukter av planter, og tiltrekker seg pollinatorer og spredere av frø og frukt.
Leukoplaster- Dette er hvite eller fargeløse plastider, for det meste runde eller ovale i formen. De er vanlige i ikke-fotosyntetiske deler av planter, som bladskinn, potetknoller osv. De lagrer næringsstoffer, oftest stivelse, men hos noen planter kan det være proteiner eller olje.
Plastider dannes i planteceller fra proplastider, som allerede er tilstede i cellene i utdanningsvevet og er små to-membranlegemer. På de tidlige stadiene av utviklingen er forskjellige typer plastider i stand til å bli til hverandre: når de utsettes for lys, blir leukoplastene til en potetknoll og kromoplastene til en gulrotrot grønne.
Plastider og mitokondrier kalles semi-autonome celleorganeller, siden de har egne DNA-molekyler og ribosomer, utfører proteinsyntese og deler seg uavhengig av celledeling. Disse funksjonene forklares av opprinnelsen fra encellede prokaryote organismer. Imidlertid er "uavhengigheten" til mitokondrier og plastider begrenset, siden deres DNA inneholder for få gener for fri eksistens, mens resten av informasjonen er kodet i kromosomene til kjernen, som gjør at den kan kontrollere disse organellene.
Endoplasmatisk retikulum (ER), eller endoplasmatisk retikulum (ER), er en enkeltmembranorganell, som er et nettverk av membranhulrom og tubuli, som okkuperer opptil 30% av innholdet i cytoplasmaet. Diameteren til ER-tubuli er omtrent 25–30 nm. Det finnes to typer EPS - grov og glatt. Grov XPS bærer ribosomer og er der proteiner syntetiseres. Glatt EPS uten ribosomer. Dens funksjon er syntese av lipider og karbohydrater, samt transport, lagring og avhending av giftige stoffer. Det er spesielt utviklet i de cellene hvor intensive metabolske prosesser finner sted, for eksempel i leverceller - hepatocytter - og skjelettmuskelfibre. Stoffer syntetisert i EPS transporteres til Golgi-apparatet. I ER blir også cellemembraner satt sammen, men dannelsen fullføres i Golgi-apparatet.
Golgi-apparatet, eller golgi kompleks, er en enkeltmembranorganell dannet av et system av flate sisterner, tubuli og vesikler snøret av dem. Den strukturelle enheten til Golgi-apparatet er diktyosome- en stabel med tanker, på den ene polen hvor stoffer fra ER kommer, og fra den motsatte polen, etter å ha gjennomgått visse transformasjoner, blir de pakket inn i bobler og sendt til andre deler av cellen. Diameteren på tankene er omtrent 2 mikron, og små bobler er omtrent 20-30 mikron. Hovedfunksjonene til Golgi-komplekset er syntesen av visse stoffer og modifisering (endring) av proteiner, lipider og karbohydrater som kommer fra ER, den endelige dannelsen av membraner, samt transport av stoffer gjennom cellen, fornyelse av dens strukturer og dannelsen av lysosomer. Golgi-apparatet fikk navnet sitt til ære for den italienske forskeren Camillo Golgi, som først oppdaget denne organoiden (1898).

Lysosomer- små enkeltmembranorganeller opp til 1 mikron i diameter, som inneholder hydrolytiske enzymer involvert i intracellulær fordøyelse. Membranene til lysosomer er dårlig permeable for disse enzymene, så ytelsen til deres funksjoner av lysosomer er veldig nøyaktig og målrettet. Så de tar en aktiv del i prosessen med fagocytose, danner fordøyelsesvakuoler, og i tilfelle sult eller skade på visse deler av cellen, fordøyer de dem uten å påvirke andre. Nylig har lysosomes rolle i celledødsprosesser blitt oppdaget.
Vakuol- et hulrom i cytoplasmaet til plante- og dyreceller, avgrenset av en membran og fylt med væske. Fordøyelses- og kontraktile vakuoler finnes i protozoceller. Førstnevnte deltar i prosessen med fagocytose, da de bryter ned næringsstoffer. Sistnevnte sikrer opprettholdelse av vann-saltbalanse på grunn av osmoregulering. Hos flercellede dyr finnes hovedsakelig fordøyelsesvakuoler.
I planteceller er vakuoler alltid til stede, de er omgitt av en spesiell membran og fylt med cellesaft. Membranen som omgir vakuolen er lik plasmamembranen i kjemisk sammensetning, struktur og funksjoner. cellesaft representerer en vandig løsning av ulike uorganiske og organiske stoffer, inkludert mineralsalter, organiske syrer, karbohydrater, proteiner, glykosider, alkaloider osv. Vakuolen kan okkupere opptil 90 % av cellevolumet og skyve kjernen til periferien. Denne delen av cellen utfører lagring, utskillelse, osmotiske, beskyttende, lysosomale og andre funksjoner, siden den akkumulerer næringsstoffer og avfallsprodukter, den gir vannforsyning og opprettholder cellens form og volum, og inneholder også enzymer for nedbrytning av mange cellekomponenter. I tillegg kan de biologisk aktive stoffene i vakuoler hindre mange dyr i å spise disse plantene. I en rekke planter, på grunn av hevelse av vakuoler, skjer cellevekst ved strekking.
Vakuoler er også tilstede i cellene til noen sopp og bakterier, men i sopp utfører de bare funksjonen osmoregulering, mens de i cyanobakterier opprettholder oppdrift og deltar i prosessene med nitrogenopptak fra luften.
Ribosomer- små ikke-membranorganeller med en diameter på 15-20 mikron, bestående av to underenheter - store og små. Eukaryote ribosomunderenheter settes sammen i nukleolen og transporteres deretter til cytoplasmaet. Ribosomene til prokaryoter, mitokondrier og plastider er mindre enn eukaryotene. Ribosomunderenheter inkluderer rRNA og proteiner.

Antall ribosomer i en celle kan nå flere titalls millioner: i cytoplasma, mitokondrier og plastider er de i fri tilstand, og på den grove ER er de i bundet tilstand. De deltar i proteinsyntese, spesielt de utfører translasjonsprosessen - biosyntesen av en polypeptidkjede på et mRNA-molekyl. På frie ribosomer syntetiseres proteiner av hyaloplasma, mitokondrier, plastider og egne proteiner av ribosomer, mens på ribosomer festet til den grove ER blir proteiner oversatt for utskillelse fra celler, montering av membraner, dannelse av lysosomer og vakuoler.
Ribosomer kan lokaliseres i hyaloplasma enkeltvis eller satt sammen i grupper med samtidig syntese av flere polypeptidkjeder på ett mRNA. Disse gruppene av ribosomer kalles polyribosomer, eller polysomer.

mikrotubuli– Dette er sylindriske hule ikke-membranorganeller som trenger inn i hele cytoplasmaet i cellen. Deres diameter er omtrent 25 nm, veggtykkelsen er 6-8 nm. De består av mange proteinmolekyler. tubulin, som først danner 13 tråder som ligner perler og deretter settes sammen til en mikrotubuli. Mikrotubuli danner et cytoplasmatisk retikulum som gir cellen form og volum, forbinder plasmamembranen med andre deler av cellen, sørger for transport av stoffer gjennom cellen, deltar i bevegelsen av cellen og intracellulære komponenter, samt i delingen av genetisk materiale. De er en del av cellesenteret og bevegelsesorganeller - flageller og flimmerhår.
mikrofilamenter, eller mikrofilamenter, er også ikke-membranorganeller, men de har en filamentøs form og dannes ikke av tubulin, men aktinom. De deltar i prosessene med membrantransport, intercellulær gjenkjennelse, deling av cellecytoplasma og i dens bevegelse. I muskelceller gir samspillet mellom aktinmikrofilamenter og myosinfilamenter sammentrekning.
Mikrotubuli og mikrofilamenter danner det indre skjelettet i cellen cytoskjelett. Det er et komplekst nettverk av fibre som gir mekanisk støtte til plasmamembranen, bestemmer cellens form, plasseringen av cellulære organeller og deres bevegelse under celledeling.

Cellesenter- ikke-membran organell lokalisert i dyreceller nær kjernen; det er fraværende i planteceller. Lengden er omtrent 0,2–0,3 µm, og diameteren er 0,1–0,15 µm. Cellesenteret består av to sentrioler liggende i innbyrdes vinkelrette plan, og strålende sfære fra mikrotubuli. Hver sentriole er dannet av ni grupper av mikrotubuli, samlet i tre, dvs. tripletter. Cellesenteret deltar i monteringen av mikrotubuli, delingen av cellens arvelige materiale, samt i dannelsen av flageller og flimmerhår.

Bevegelsesorganeller. Flagella og flimmerhår er utvekster av celler dekket med plasmalemma. Disse organellene er basert på ni par mikrotubuli plassert langs periferien og to frie mikrotubuli i midten. Mikrotubuli er sammenkoblet av ulike proteiner som sikrer deres koordinerte avvik fra aksen - oscillasjon. Svingninger er energiavhengige, det vil si at energien til makroerge bindinger av ATP brukes på denne prosessen. Restaurering av tapte flageller og flimmerhår er en funksjon basale kropper, eller kinetosomer plassert ved basen deres.
Lengden på flimmerhårene er omtrent 10-15 nm, og lengden på flagellene er 20-50 mikron. På grunn av de strengt rettede bevegelsene til flagellene og flimmerhårene, utføres ikke bare bevegelsen av encellede dyr, sædceller, etc., men også luftveiene ryddes, egget beveger seg gjennom egglederne, siden alle disse delene av mennesket kroppen er foret med ciliert epitel.

Inkluderinger
Inkluderinger– Dette er ikke-permanente komponenter i cellen, som dannes og forsvinner i løpet av livet. Disse inkluderer både reservestoffer, for eksempel stivelses- eller proteinkorn i planteceller, glykogengranulat i dyre- og soppceller, volutin i bakterier, fettdråper i alle celletyper og avfallsprodukter, spesielt ufordøyde matrester som følge av dette. av fagocytose. , som danner de såkalte gjenværende legemer.

Forholdet mellom strukturen og funksjonene til delene og organellene i cellen er grunnlaget for dens integritet
Hver av delene av cellen er på den ene siden en egen struktur med en spesifikk struktur og funksjoner, og på den andre siden en komponent i et mer komplekst system kalt en celle. Mesteparten av den arvelige informasjonen til en eukaryot celle er konsentrert i kjernen, men selve kjernen er ikke i stand til å sikre implementeringen, siden dette krever i det minste cytoplasmaet, som fungerer som hovedstoffet, og ribosomer, som denne syntesen skjer på . De fleste ribosomer er lokalisert på det granulære endoplasmatiske retikulum, hvorfra proteiner oftest transporteres til Golgi-komplekset, og deretter, etter modifikasjon, til de delene av cellen de er beregnet på, eller skilles ut. Membranpakking av proteiner og karbohydrater kan integreres i organoide membraner og den cytoplasmatiske membranen, noe som sikrer deres konstante fornyelse. Lysosomer og vakuoler, som utfører de viktigste funksjonene, er også snøret fra Golgi-komplekset. For eksempel, uten lysosomer, ville celler raskt blitt til en slags dump av avfallsmolekyler og strukturer.
Alle disse prosessene krever energi produsert av mitokondrier og, i planter, også av kloroplaster. Og selv om disse organellene er relativt autonome, siden de har sine egne DNA-molekyler, er noen av deres proteiner fortsatt kodet av kjernegenomet og syntetisert i cytoplasmaet.
Dermed er cellen en uatskillelig enhet av dens bestanddeler, som hver utfører sin egen unike funksjon.
Metabolisme og energiomdannelse er egenskaper ved levende organismer. Energi og plastisk metabolisme, deres forhold. Stadier av energimetabolisme. Fermentering og respirasjon. Fotosyntese, dens betydning, kosmisk rolle. Faser av fotosyntese. Lys og mørke reaksjoner av fotosyntese, deres forhold. Kjemosyntese. Rollen til kjemosyntetiske bakterier på jorden
Metabolisme og energiomdannelse - egenskaper til levende organismer
Cellen kan sammenlignes med en miniatyr kjemisk fabrikk hvor hundrevis og tusenvis av kjemiske reaksjoner finner sted.
Metabolisme- et sett med kjemiske transformasjoner rettet mot bevaring og selvreproduksjon av biologiske systemer.
Det inkluderer inntak av stoffer i kroppen under ernæring og respirasjon, intracellulær metabolisme, eller metabolisme, samt tildeling av sluttprodukter av metabolisme.
Metabolisme er uløselig knyttet til prosessene med å konvertere en type energi til en annen. For eksempel, i prosessen med fotosyntese, lagres lysenergi i form av energien til kjemiske bindinger til komplekse organiske molekyler, og i prosessen med respirasjon frigjøres den og brukes på syntese av nye molekyler, mekanisk og osmotisk arbeid, spres i form av varme osv.
Flyten av kjemiske reaksjoner i levende organismer sikres av biologiske katalysatorer av proteinnatur - enzymer, eller enzymer. Som andre katalysatorer, akselererer enzymer flyten av kjemiske reaksjoner i cellen med titalls og hundretusenvis av ganger, og noen ganger til og med gjør dem mulige, men endrer verken arten eller egenskapene til sluttproduktet (produktene) av reaksjonen og ikke endre seg selv. Enzymer kan være både enkle og komplekse proteiner, som i tillegg til proteindelen også inkluderer en ikke-proteindel - kofaktor (koenzym). Eksempler på enzymer er spyttamylase, som bryter ned polysakkarider ved langvarig tygging, og pepsin, som sørger for fordøyelsen av proteiner i magen.
Enzymer skiller seg fra ikke-proteinkatalysatorer i deres høye virkningsspesifisitet, en betydelig økning i reaksjonshastigheten med deres hjelp, samt evnen til å regulere virkningen ved å endre reaksjonsbetingelsene eller interagere med forskjellige stoffer. I tillegg er forholdene for enzymatisk katalyse vesentlig forskjellig fra de som ikke-enzymatisk katalyse skjer under: temperaturen på $37°C$ er optimal for funksjonen til enzymer i menneskekroppen, trykket bør være nær atmosfærisk, og $pH$ til mediet kan nøle betydelig. Så for amylase er et alkalisk miljø nødvendig, og for pepsin et surt.
Virkningsmekanismen til enzymer er å redusere aktiveringsenergien til stoffer (substrater) som kommer inn i reaksjonen på grunn av dannelsen av mellomliggende enzym-substratkomplekser.

Energi og plastisk metabolisme, deres forhold
Metabolisme består av to prosesser som skjer samtidig i cellen: plast og energiutveksling.
Plastmetabolisme (anabolisme, assimilering) er et sett med syntesereaksjoner som følger med forbruket av ATP-energi. I prosessen med plastisk metabolisme syntetiseres organiske stoffer som er nødvendige for cellen. Eksempler på plastiske utvekslingsreaksjoner er fotosyntese, proteinbiosyntese og DNA-replikasjon (selvdobling).
Energimetabolisme (katabolisme, dissimilering) er et sett med reaksjoner som bryter ned komplekse stoffer til enklere. Som et resultat av energimetabolisme frigjøres energi, lagret i form av ATP. De viktigste prosessene for energimetabolisme er respirasjon og gjæring.
Plast- og energiutveksling er uløselig forbundet, siden i prosessen med plastutveksling syntetiseres organiske stoffer og dette krever energien til ATP, og i prosessen med energimetabolisme splittes organiske stoffer og energi frigjøres, som deretter vil bli brukt på syntese prosesser.
Organismer mottar energi i prosessen med ernæring, og frigjør den og konverterer den til en tilgjengelig form hovedsakelig i prosessen med respirasjon. I henhold til ernæringsmåten er alle organismer delt inn i autotrofer og heterotrofer. Autotrofer i stand til selvstendig å syntetisere organiske stoffer fra uorganiske, og heterotrofer bruk utelukkende ferdige organiske stoffer.
Stadier av energimetabolisme
Til tross for kompleksiteten til energimetabolismereaksjoner, er den betinget delt inn i tre stadier: forberedende, anaerob (oksygenfri) og aerob (oksygen).
På forberedende stadium molekyler av polysakkarider, lipider, proteiner, nukleinsyrer brytes ned til enklere, for eksempel glukose, glyserol og fettsyrer, aminosyrer, nukleotider osv. Dette stadiet kan finne sted direkte i cellene eller i tarmen, hvorfra splittede stoffer leveres med blodstrøm.
anaerobt stadium energimetabolismen er ledsaget av ytterligere spaltning av monomerene til organiske forbindelser til enda enklere mellomprodukter, for eksempel pyrodruesyre eller pyruvat. Det krever ikke tilstedeværelse av oksygen, og for mange organismer som lever i sumpenes silt eller i menneskets tarm, er det den eneste måten å få energi på. Det anaerobe stadiet av energimetabolismen finner sted i cytoplasmaet.
Ulike stoffer kan gjennomgå oksygenfri spaltning, men glukose er ofte substratet for reaksjonene. Prosessen med oksygenfri spaltning kalles glykolyse. Under glykolyse mister glukosemolekylet fire hydrogenatomer, det vil si at det oksideres, og to molekyler pyrodruesyre, to ATP-molekyler og to molekyler av den reduserte hydrogenbæreren $NADH + H^(+)$ dannes:
$C_6H_(12)O_6 + 2H_3PO_4 + 2ADP + 2NAD → 2C_3H_4O_3 + 2ATP + 2NADH + H^(+) + 2H_2O$.
Dannelsen av ATP fra ADP skjer på grunn av direkte overføring av et fosfatanion fra et tidligere fosforylert sukker og kalles substratfosforylering.
Aerobic scene energiutveksling kan bare skje i nærvær av oksygen, mens mellomforbindelsene som dannes i prosessen med oksygenfri spaltning oksideres til sluttprodukter (karbondioksid og vann) og mesteparten av energien som er lagret i de kjemiske bindingene til organiske forbindelser frigjøres . Det går over i energien til makroerge bindinger av 36 ATP-molekyler. Dette stadiet kalles også vevsånding. I fravær av oksygen omdannes mellomforbindelser til andre organiske stoffer, en prosess som kalles gjæring.
Pust
Mekanismen for cellulær respirasjon er skjematisk vist i fig.

Aerob respirasjon forekommer i mitokondrier, mens pyrodruesyre først mister ett karbonatom, som er ledsaget av syntesen av en reduserende ekvivalent av $NADH + H^(+)$ og et acetylkoenzym A (acetyl-CoA)-molekyl:
$C_3H_4O_3 + NAD + H~CoA → CH_3CO~CoA + NADH + H^(+) + CO_2$.
Acetyl-CoA i mitokondriematrisen er involvert i en kjede av kjemiske reaksjoner, hvis helhet kalles Krebs syklus (trikarboksylsyresyklus, sitronsyresyklus). Under disse transformasjonene dannes det to ATP-molekyler, acetyl-CoA oksideres fullstendig til karbondioksid, og dets hydrogenioner og elektroner er festet til hydrogenbærerne $NADH + H^(+)$ og $FADH_2$. Bærere transporterer hydrogenprotoner og elektroner til de indre membranene i mitokondriene, som danner cristae. Ved hjelp av bærerproteiner pumpes hydrogenprotoner inn i intermembranrommet, og elektroner overføres langs den såkalte respiratoriske kjeden av enzymer som ligger på den indre membranen av mitokondrier og dumpes på oksygenatomer:
$O_2+2e^(-)→O_2^-$.
Det skal bemerkes at noen proteiner i respirasjonskjeden inneholder jern og svovel.
Fra intermembranrommet transporteres hydrogenprotoner tilbake til mitokondriematrisen ved hjelp av spesielle enzymer - ATP-syntaser, og energien som frigjøres i dette tilfellet brukes på syntese av 34 ATP-molekyler fra hvert glukosemolekyl. Denne prosessen kalles oksidativ fosforylering. I mitokondriematrisen reagerer hydrogenprotoner med oksygenradikaler for å danne vann:
$4H^(+)+O_2^-→2H_2O$.
Settet med reaksjoner av oksygenrespirasjon kan uttrykkes som følger:
$2C_3H_4O_3 + 6O_2 + 36H_3PO_4 + 36ADP → 6CO_2 + 38H_2O + 36ATP.$
Den generelle pusteligningen ser slik ut:
$C_6H_(12)O_6 + 6O_2 + 38H_3PO_4 + 38ADP → 6CO_2 + 40H_2O + 38ATP.$
Fermentering
I fravær av oksygen eller mangel på det, oppstår gjæring. Fermentering er en evolusjonært tidligere måte å få energi på enn respirasjon, men det er energimessig mindre lønnsomt, siden fermentering produserer organiske stoffer som fortsatt er rike på energi. Det er flere hovedtyper av gjæring: melkesyre, alkohol, eddiksyre, etc. Så i skjelettmuskulatur, i fravær av oksygen under gjæring, reduseres pyrodruesyre til melkesyre, mens de tidligere dannede reduserende ekvivalentene forbrukes, og bare to ATP-molekyler gjenstår:
$2C_3H_4O_3 + 2NADH + H^(+) → 2C_3H_6O_3 + 2NAD$.
Under gjæring ved hjelp av gjærsopp blir pyrodruesyre i nærvær av oksygen til etylalkohol og karbonmonoksid (IV):
$C_3H_4O_3 + NADH + H^(+) → C_2H_5OH + CO_2 + NAD^(+)$.
Under fermentering ved hjelp av mikroorganismer kan pyrodruesyre også danne eddiksyre, smørsyre, maursyre, etc.
ATP oppnådd som et resultat av energimetabolisme forbrukes i cellen for ulike typer arbeid: kjemisk, osmotisk, elektrisk, mekanisk og regulatorisk. Kjemisk arbeid består i biosyntese av proteiner, lipider, karbohydrater, nukleinsyrer og andre vitale forbindelser. Osmotisk arbeid inkluderer prosessene med absorpsjon av cellen og fjerning fra den av stoffer som er i det ekstracellulære rommet i konsentrasjoner større enn i selve cellen. Elektrisk arbeid er nært knyttet til osmotisk arbeid, siden det er som et resultat av bevegelsen av ladede partikler gjennom membranene at ladningen til membranen dannes og egenskapene til eksitabilitet og ledningsevne erverves. Mekanisk arbeid er assosiert med bevegelse av stoffer og strukturer inne i cellen, så vel som cellen som helhet. Reguleringsarbeid omfatter alle prosesser som tar sikte på å koordinere prosesser i cellen.
Fotosyntese, dens betydning, kosmisk rolle
fotosyntese kalt prosessen med å konvertere lysenergi til energien til kjemiske bindinger av organiske forbindelser med deltakelse av klorofyll.
Som et resultat av fotosyntesen produseres det årlig rundt 150 milliarder tonn organisk materiale og omtrent 200 milliarder tonn oksygen. Denne prosessen sikrer sirkulasjon av karbon i biosfæren, forhindrer akkumulering av karbondioksid og forhindrer derved forekomsten av drivhuseffekten og overoppheting av jorden. De organiske stoffene som dannes som følge av fotosyntesen blir ikke fullstendig konsumert av andre organismer, en betydelig del av dem dannet mineralforekomster (hardt og brunt kull, olje) over millioner av år. Nylig har også rapsolje («biodiesel») og alkohol hentet fra planterester blitt brukt som drivstoff. Fra oksygen, under påvirkning av elektriske utladninger, dannes ozon, som danner et ozonskjold som beskytter alt liv på jorden mot de skadelige effektene av ultrafiolette stråler.
Vår landsmann, den fremragende plantefysiologen K. A. Timiryazev (1843-1920) kalte rollen til fotosyntesen "kosmisk", siden den forbinder jorden med solen (rommet), og gir en tilstrømning av energi til planeten.
Faser av fotosyntese. Lys og mørke reaksjoner av fotosyntese, deres forhold
I 1905 oppdaget den engelske plantefysiologen F. Blackman at fotosyntesehastigheten ikke kan øke i det uendelige, en eller annen faktor begrenser den. Basert på dette foreslo han eksistensen av to faser av fotosyntese: lys og mørk. Ved lav lysintensitet øker lysreaksjonshastigheten proporsjonalt med økningen i lysintensitet, og i tillegg er disse reaksjonene ikke avhengig av temperatur, siden enzymer ikke er nødvendige for deres forekomst. Lysreaksjoner forekommer på thylakoidmembraner.
Hastigheten av mørkereaksjoner øker tvert imot med økende temperatur, men når temperaturterskelen på $30 °C $ nås, stopper denne veksten, noe som indikerer den enzymatiske naturen til disse transformasjonene som skjer i stroma. Det skal bemerkes at lys også har en viss effekt på mørkereaksjoner, til tross for at de kalles mørke.

Den lette fasen av fotosyntesen fortsetter på thylakoidmembraner, som bærer flere typer proteinkomplekser, hvorav de viktigste er fotosystemer I og II, samt ATP-syntase. Sammensetningen av fotosystemer inkluderer pigmentkomplekser, der det i tillegg til klorofyll også er karotenoider. Karotenoider fanger lys i de områdene av spekteret der klorofyll ikke gjør det, og beskytter også klorofyll mot ødeleggelse av lys med høy intensitet.
I tillegg til pigmentkomplekser inkluderer fotosystemer også en rekke elektronakseptorproteiner som suksessivt overfører elektroner fra klorofyllmolekyler til hverandre. Rekkefølgen til disse proteinene kalles kloroplast elektron transportkjede.
Et spesielt kompleks av proteiner er også assosiert med fotosystem II, som sikrer frigjøring av oksygen under fotosyntesen. Dette oksygenutviklende komplekset inneholder mangan- og klorioner.
PÅ lett fase lyskvanter, eller fotoner, som faller på klorofyllmolekyler som ligger på thylakoidmembraner, overfører dem til en eksitert tilstand preget av høyere elektronenergi. Samtidig overføres eksiterte elektroner fra klorofyllet til fotosystem I gjennom en kjede av mellomledd til hydrogenbæreren NADP, som tilfører hydrogenprotoner, som alltid er tilstede i en vandig løsning:
$NADP + 2e^(-) + 2H^(+) → NADPH + H^(+)$.
Den reduserte $NADPH + H^(+)$ vil deretter bli brukt i det mørke stadiet. Elektroner fra klorofyllet til fotosystem II overføres også langs elektrontransportkjeden, men de fyller "elektronhullene" i klorofyllet til fotosystem I. Mangelen på elektroner i klorofyllet til fotosystem II fylles ved å frata vannmolekyler vannmolekyler , som forekommer med deltakelse av det oksygenfrigjørende komplekset som allerede er nevnt ovenfor. Som et resultat av nedbrytning av vannmolekyler, som kalles fotolyse, dannes hydrogenprotoner og molekylært oksygen frigjøres, som er et biprodukt av fotosyntesen:
$H_2O → 2H^(+) + 2e^(-) + (1)/(2)O_2$.
Genetisk informasjon i en celle. Gener, genetisk kode og dens egenskaper. Matrisearten til biosyntetiske reaksjoner. Biosyntese av protein og nukleinsyrer
Genetisk informasjon i en celle
Reproduksjon av ens egen art er en av de grunnleggende egenskapene til de levende. På grunn av dette fenomenet er det en likhet ikke bare mellom organismer, men også mellom individuelle celler, så vel som deres organeller (mitokondrier og plastider). Det materielle grunnlaget for denne likheten er overføring av genetisk informasjon kryptert i DNA-nukleotidsekvensen, som utføres på grunn av prosessene med DNA-replikasjon (selvdobling). Alle funksjoner og egenskaper til celler og organismer realiseres takket være proteiner, hvis struktur først og fremst bestemmes av DNA-nukleotidsekvensene. Derfor er det biosyntesen av nukleinsyrer og proteiner som er av største betydning i metabolske prosesser. Den strukturelle enheten for arvelig informasjon er genet.
Gener, genetisk kode og dens egenskaper
Arvelig informasjon i en celle er ikke monolitisk, den er delt inn i separate "ord" - gener.
Gene er den grunnleggende enheten for genetisk informasjon.
Arbeidet med "Human Genome"-programmet, som ble utført samtidig i flere land og ble fullført på begynnelsen av dette århundret, ga oss en forståelse av at en person bare har rundt 25-30 tusen gener, men informasjon fra de fleste av våre DNA leses aldri, siden det inneholder et stort antall meningsløse seksjoner, repetisjoner og gener som koder for funksjoner som har mistet sin betydning for mennesker (hale, kroppshår osv.). I tillegg er en rekke gener som er ansvarlige for utviklingen av arvelige sykdommer, samt medikamentmålgener, blitt dechiffrert. Den praktiske anvendelsen av resultatene som er oppnådd under implementeringen av dette programmet blir imidlertid utsatt til genomene til flere mennesker er dekodet og det blir klart hvordan de skiller seg.
Gener som koder for den primære strukturen til et protein, ribosomalt eller overførings-RNA kalles strukturell, og gener som gir aktivering eller undertrykkelse av lesing av informasjon fra strukturelle gener - regulatoriske. Imidlertid inneholder selv strukturelle gener regulatoriske regioner.
Den arvelige informasjonen til organismer er kryptert i DNA i form av visse kombinasjoner av nukleotider og deres sekvens - genetisk kode. Dens egenskaper er: triplett, spesifisitet, universalitet, redundans og ikke-overlappende. I tillegg er det ingen skilletegn i den genetiske koden.
Hver aminosyre er kodet i DNA av tre nukleotider. trilling for eksempel er metionin kodet av TAC-tripletten, det vil si triplettkoden. På den annen side koder hver triplett bare for én aminosyre, som er dens spesifisitet eller entydighet. Den genetiske koden er universell for alle levende organismer, det vil si at arvelig informasjon om menneskelige proteiner kan leses av bakterier og omvendt. Dette vitner om enheten i opprinnelsen til den organiske verden. Imidlertid tilsvarer 64 kombinasjoner av tre nukleotider kun 20 aminosyrer, som et resultat av hvilke 2-6 tripletter kan kode for én aminosyre, det vil si at den genetiske koden er overflødig, eller degenerert. Tre trillinger har ikke tilsvarende aminosyrer, kalles de stopp kodoner, da de markerer slutten på syntesen av polypeptidkjeden.
Sekvensen av baser i DNA-tripletter og aminosyrene de koder for

*Stoppkodon, som indikerer slutten av syntesen av polypeptidkjeden.
Forkortelser for aminosyrenavn:
Ala - alanin
Arg - arginin
Asn - asparagin
Asp - asparaginsyre
Val - valine
Hans - histidin
Gly - glycin
Gln - glutamin
Glu - glutaminsyre
Ile - isoleucin
Leu - leucin
Liz - lysin
Meth - metionin
Pro - prolin
Ser - serin
Tyr - tyrosin
Tre - treonin
Tre - tryptofan
Fen - fenylalanin
cis - cystein
Hvis du begynner å lese genetisk informasjon ikke fra det første nukleotidet i tripletten, men fra det andre, vil ikke bare leserammen skifte, proteinet syntetisert på denne måten vil være helt annerledes, ikke bare i nukleotidsekvensen, men også i strukturen og eiendommer. Det er ingen skilletegn mellom trillingene, så det er ingen hindringer for forskyvning av leserammen, noe som åpner for forekomst og vedlikehold av mutasjoner.
Matrisearten til biosyntetiske reaksjoner
Bakterieceller er i stand til å duplisere hvert 20.-30. minutt, og eukaryote celler - hver dag og enda oftere, noe som krever høy hastighet og nøyaktighet av DNA-replikasjon. I tillegg inneholder hver celle hundrevis og tusenvis av kopier av mange proteiner, spesielt enzymer, derfor er "stykke"-metoden for deres produksjon uakseptabel for deres reproduksjon. En mer progressiv måte er stempling, som lar deg få mange eksakte kopier av produktet og også redusere kostnadene. For stempling er det nødvendig med en matrise, som et inntrykk blir gjort med.
I celler er prinsippet for matrisesyntese at nye molekyler av proteiner og nukleinsyrer syntetiseres i henhold til programmet som er lagt i strukturen til eksisterende molekyler av de samme nukleinsyrene (DNA eller RNA).
Biosyntese av protein og nukleinsyrer
DNA-replikasjon. DNA er en dobbelttrådet biopolymer hvis monomerer er nukleotider. Hvis DNA-biosyntesen foregikk i henhold til prinsippet om fotokopiering, ville det uunngåelig oppstå mange forvrengninger og feil i arvelig informasjon, noe som til slutt ville føre til døden til nye organismer. Derfor er prosessen med DNA-duplisering annerledes, på en semi-konservativ måte: DNA-molekylet vikler seg ut, og på hver av kjedene syntetiseres en ny kjede etter komplementaritetsprinsippet. Prosessen med selvreproduksjon av DNA-molekylet, som sikrer nøyaktig kopiering av arvelig informasjon og dens overføring fra generasjon til generasjon, kalles replikering(fra lat. replikering- repetisjon). Som et resultat av replikasjon dannes det to absolutt eksakte kopier av det overordnede DNA-molekylet, som hver har en kopi av det overordnede.

Replikasjonsprosessen er faktisk ekstremt kompleks, siden en rekke proteiner er involvert i den. Noen av dem vikler ut dobbelthelixen til DNA, andre bryter hydrogenbindingene mellom nukleotidene i komplementære kjeder, andre (for eksempel DNA-polymerase-enzymet) velger nye nukleotider etter komplementaritetsprinsippet osv. De to DNA-molekylene dannet som et resultat av replikasjon divergerer i to under deling, nydannede datterceller.
Feil i replikasjonsprosessen er ekstremt sjeldne, men hvis de oppstår, elimineres de veldig raskt av både DNA-polymeraser og spesielle reparasjonsenzymer, siden enhver feil i nukleotidsekvensen kan føre til en irreversibel endring i strukturen og funksjonene til proteinet og til slutt påvirke levedyktigheten til en ny celle eller til og med et individ negativt.
proteinbiosyntese. Som den fremragende filosofen på 1800-tallet F. Engels i overført betydning sa det: "Livet er en form for eksistens av proteinlegemer." Strukturen og egenskapene til proteinmolekyler bestemmes av deres primære struktur, dvs. sekvensen av aminosyrer kodet i DNA. Ikke bare eksistensen av selve polypeptidet, men også funksjonen til cellen som helhet avhenger av nøyaktigheten av reproduksjonen av denne informasjonen; derfor er prosessen med proteinsyntese av stor betydning. Det ser ut til å være den mest komplekse synteseprosessen i cellen, siden opptil tre hundre forskjellige enzymer og andre makromolekyler er involvert her. I tillegg flyter den med høy hastighet, noe som krever enda større presisjon.
Det er to hovedtrinn i proteinbiosyntese: transkripsjon og translasjon.
Transkripsjon(fra lat. transkripsjon- omskriving) er biosyntesen av mRNA-molekyler på en DNA-mal.

Siden DNA-molekylet inneholder to antiparallelle kjeder, vil lesing av informasjon fra begge kjeder føre til dannelse av helt forskjellige mRNA-er, derfor er biosyntesen deres mulig bare på en av kjedene, som kalles koding, eller kodogen, i motsetning til den andre, ikke-kodende eller ikke-kodogene. Omskrivingsprosessen leveres av et spesielt enzym, RNA-polymerase, som velger RNA-nukleotider i henhold til komplementaritetsprinsippet. Denne prosessen kan foregå både i kjernen og i organeller som har sitt eget DNA - mitokondrier og plastider.
mRNA-molekylene syntetisert under transkripsjon gjennomgår en kompleks prosess med forberedelse til translasjon (mitokondrie- og plastid-mRNA-er kan forbli inne i organeller, der det andre stadiet av proteinbiosyntesen finner sted). I prosessen med mRNA-modning festes de tre første nukleotidene (AUG) og en hale av adenylnukleotider til den, hvor lengden bestemmer hvor mange proteinkopier som kan syntetiseres på et gitt molekyl. Først da forlater modne mRNA-er kjernen gjennom kjernefysiske porer.
Parallelt skjer prosessen med aminosyreaktivering i cytoplasmaet, hvor aminosyren festes til det tilsvarende frie tRNA. Denne prosessen er katalysert av et spesielt enzym, det bruker ATP.
Kringkaste(fra lat. kringkaste- transfer) er biosyntesen av en polypeptidkjede på en mRNA-mal, der genetisk informasjon blir oversatt til en sekvens av aminosyrer i polypeptidkjeden.

Det andre stadiet av proteinsyntese forekommer oftest i cytoplasmaet, for eksempel på det grove endoplasmatiske retikulum. Dens forekomst krever tilstedeværelse av ribosomer, aktivering av tRNA, der de fester de tilsvarende aminosyrene, tilstedeværelsen av Mg2+ ioner, samt optimale miljøforhold (temperatur, pH, trykk, etc.).
For å begynne å kringkaste innvielse) en liten underenhet av ribosomet festes til mRNA-molekylet klar for syntese, og deretter, i henhold til komplementaritetsprinsippet, velges tRNA som bærer aminosyren metionin til det første kodonet (AUG). Først da slutter den store underenheten til ribosomet seg. Innenfor det sammensatte ribosomet er det to mRNA-kodoner, hvorav det første allerede er okkupert. Et andre tRNA, som også bærer en aminosyre, festes til kodonet ved siden av det, hvoretter det dannes en peptidbinding mellom aminosyrerestene ved hjelp av enzymer. Ribosomet flytter ett kodon av mRNA; det første av tRNA, frigjort fra aminosyren, går tilbake til cytoplasmaet for den neste aminosyren, og et fragment av den fremtidige polypeptidkjeden henger så å si på det gjenværende tRNA. Det neste tRNA slutter seg til det nye kodonet, som er innenfor ribosomet, prosessen gjentas og trinn for trinn forlenges polypeptidkjeden, dvs. forlengelse.
Slutt på proteinsyntesen avslutning) oppstår så snart en spesifikk nukleotidsekvens påtreffes i et mRNA-molekyl som ikke koder for en aminosyre (stoppkodon). Deretter separeres ribosomet, mRNA og polypeptidkjeden, og det nysyntetiserte proteinet får den passende strukturen og transporteres til den delen av cellen hvor det skal utføre sine funksjoner.
Translasjon er en svært energikrevende prosess, siden energien til ett ATP-molekyl brukes på å feste én aminosyre til tRNA, og flere brukes til å flytte ribosomet langs mRNA-molekylet.
For å akselerere syntesen av visse proteinmolekyler, kan flere ribosomer festes sekvensielt til mRNA-molekylet, som danner en enkelt struktur - polysom.
Cellen er den genetiske enheten til levende ting. Kromosomer, deres struktur (form og størrelse) og funksjoner. Antall kromosomer og deres artskonstans. Somatiske og kjønnsceller. Cellelivssyklus: interfase og mitose. Mitose er delingen av somatiske celler. Meiose. Faser av mitose og meiose. Utviklingen av kjønnsceller i planter og dyr. Celledeling er grunnlaget for vekst, utvikling og reproduksjon av organismer. Rollen til meiose og mitose
Cellen er livets genetiske enhet
Til tross for det faktum at nukleinsyrer er bæreren av genetisk informasjon, er implementeringen av denne informasjonen umulig utenfor cellen, noe som lett kan bevises ved eksemplet med virus. Disse organismene, som ofte bare inneholder DNA eller RNA, kan ikke formere seg på egen hånd, for dette må de bruke cellens arvelige apparat. De kan ikke engang trenge inn i cellen uten hjelp fra cellen selv, bortsett fra ved å bruke mekanismene for membrantransport eller på grunn av celleskade. De fleste virus er ustabile, de dør etter noen timers eksponering for friluft. Derfor er cellen en genetisk enhet av de levende, som har et minimumssett med komponenter for bevaring, modifikasjon og implementering av arvelig informasjon, samt overføring til etterkommere.
Mesteparten av den genetiske informasjonen til en eukaryot celle er lokalisert i kjernen. Et trekk ved organisasjonen er at, i motsetning til DNAet til en prokaryot celle, er ikke eukaryote DNA-molekyler lukket og danner komplekse komplekser med proteiner - kromosomer.
Kromosomer, deres struktur (form og størrelse) og funksjoner
Kromosom(fra gresk. krom- farge, farge og steinbit- kropp) er strukturen til cellekjernen, som inneholder gener og bærer viss arvelig informasjon om kroppens tegn og egenskaper.
Noen ganger kalles ring-DNA-molekylene til prokaryoter også kromosomer. Kromosomer er i stand til selvduplisering, de har en strukturell og funksjonell individualitet og beholder den i flere generasjoner. Hver celle bærer all arvelig informasjon fra kroppen, men bare en liten del av den fungerer.
Grunnlaget for kromosomet er et dobbelttrådet DNA-molekyl fullpakket med proteiner. Hos eukaryoter samhandler histon- og ikke-histonproteiner med DNA, mens histonproteiner er fraværende i prokaryoter.
Kromosomer ses best under et lysmikroskop under celledeling, når de, som et resultat av komprimering, tar form av stavformede kropper adskilt av en primær innsnevring - sentromer — på skuldrene. Kromosomet kan også ha sekundær innsnevring, som i noen tilfeller skiller den såkalte satellitt. Endene av kromosomene kalles telomerer. Telomerer hindrer endene av kromosomene i å feste seg sammen og sikrer at de fester seg til kjernemembranen i en celle som ikke deler seg. Ved begynnelsen av deling dobles kromosomene og består av to datterkromosomer - kromatider festet ved sentromeren.

I henhold til formen skilles kromosomer med like arm, ulik arm og stavformede kromosomer. Kromosomstørrelsene varierer betydelig, men det gjennomsnittlige kromosomet har en størrelse på 5 $×$ 1,4 µm.
I noen tilfeller inneholder kromosomer, som et resultat av tallrike DNA-duplikasjoner, hundrevis og tusenvis av kromatider: slike gigantiske kromosomer kalles polyeten. De finnes i spyttkjertlene til Drosophila-larvene, så vel som i fordøyelseskjertlene til rundorm.
Antall kromosomer og deres artskonstans. Somatiske og kjønnsceller
I følge den cellulære teorien er en celle en enhet av struktur, liv og utvikling av en organisme. Dermed er slike viktige funksjoner av levende ting som vekst, reproduksjon og utvikling av organismen gitt på cellenivå. Celler fra flercellede organismer kan deles inn i somatisk og kjønn.
somatiske celler er alle cellene i kroppen som er dannet som et resultat av mitotisk deling.
Studiet av kromosomer gjorde det mulig å fastslå at de somatiske cellene i organismen til hver biologisk art er preget av et konstant antall kromosomer. For eksempel har en person 46. Settet med kromosomer av somatiske celler kalles diploid(2n), eller dobbel.

kjønnsceller, eller kjønnsceller, er spesialiserte celler som tjener til seksuell reproduksjon.
Gameter inneholder alltid halvparten så mange kromosomer som i somatiske celler (hos mennesker - 23), så settet med kromosomer til kjønnsceller kalles haploid(n), eller singel. Dannelsen er assosiert med meiotisk celledeling.
Mengden DNA fra somatiske celler er betegnet som 2c, og mengden av kjønnsceller er 1c. Den genetiske formelen til somatiske celler er skrevet som 2n2c, og kjønn - 1n1c.
I kjernene til noen somatiske celler kan antallet kromosomer avvike fra antallet i somatiske celler. Hvis denne forskjellen er større med ett, to, tre, etc. haploide sett, kalles slike celler polyploid(henholdsvis tri-, tetra-, pentaploid). I slike celler er metabolske prosesser vanligvis svært intensive.
Antall kromosomer i seg selv er ikke en artsspesifikk egenskap, siden ulike organismer kan ha samme antall kromosomer, mens beslektede kan ha forskjellig antall. For eksempel har malariaplasmodium og hesterundorm to kromosomer, mens mennesker og sjimpanser har henholdsvis 46 og 48.
Menneskelige kromosomer er delt inn i to grupper: autosomer og kjønnskromosomer (heterokromosomer). Autosome det er 22 par i menneskelige somatiske celler, de er like for menn og kvinner, og kjønnskromosomer bare ett par, men det er hun som bestemmer kjønnet til individet. Det er to typer kjønnskromosomer - X og Y. Cellene i kroppen til en kvinne bærer to X-kromosomer, og menn - X og Y.
Karyotype- dette er et sett med tegn på kromosomsettet til en organisme (antall kromosomer, deres form og størrelse).
Den betingede registreringen av karyotypen inkluderer det totale antallet kromosomer, kjønnskromosomer og mulige avvik i settet med kromosomer. For eksempel er karyotypen til en normal mann skrevet som 46,XY, mens karyotypen til en normal kvinne er 46,XX.
Cellelivssyklus: interfase og mitose
Celler oppstår ikke hver gang på nytt, de dannes kun som et resultat av deling av moderceller. Etter separasjon tar dattercellene litt tid på å danne organeller og skaffe seg den passende strukturen som vil sikre utførelsen av en viss funksjon. Denne tidsperioden kalles modning.
Tidsperioden fra en celle dukker opp som et resultat av deling til dens deling eller død kalles celle livssyklus.
I eukaryote celler er livssyklusen delt inn i to hovedstadier: interfase og mitose.
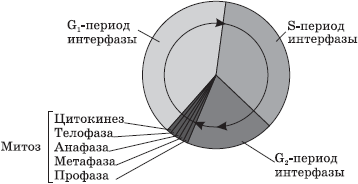
Interfase- dette er tidsperioden i livssyklusen hvor cellen ikke deler seg og fungerer normalt. Interfasen er delt inn i tre perioder: G 1 -, S- og G 2 -perioder.
G 1 -periode(presyntetisk, postmitotisk) er en periode med cellevekst og utvikling, der det er en aktiv syntese av RNA, proteiner og andre stoffer som er nødvendige for fullstendig livsstøtte til den nydannede cellen. Ved slutten av denne perioden kan cellen begynne å forberede seg på DNA-duplisering.
PÅ S-periode(syntetisk) prosessen med DNA-replikasjon finner sted. Den eneste delen av kromosomet som ikke gjennomgår replikasjon er sentromeren, derfor divergerer de resulterende DNA-molekylene ikke helt, men forblir festet i det, og i begynnelsen av delingen har kromosomet et X-formet utseende. Den genetiske formelen til cellen etter DNA-duplisering er 2n4c. Også i S-perioden skjer dobling av centriolene til cellesenteret.
G 2 -periode(postsyntetisk, premitotisk) er preget av intensiv syntese av RNA, proteiner og ATP som er nødvendig for prosessen med celledeling, samt separasjon av sentrioler, mitokondrier og plastider. Inntil slutten av interfasen forblir kromatinet og nukleolus tydelig skillelig, integriteten til kjernemembranen blir ikke forstyrret, og organellene endres ikke.
Noen av kroppens celler er i stand til å utføre sine funksjoner gjennom hele kroppens liv (nevroner i hjernen vår, muskelceller i hjertet), mens andre eksisterer i kort tid, hvoretter de dør (celler i tarmepitelet). , celler i hudens epidermis). Følgelig må prosessene med celledeling og dannelsen av nye celler hele tiden skje i kroppen, som ville erstatte de døde. Celler som er i stand til å dele kalles stilk. I menneskekroppen finnes de i den røde benmargen, i de dype lagene av hudens epidermis og andre steder. Ved å bruke disse cellene kan du dyrke et nytt organ, oppnå foryngelse og også klone kroppen. Utsiktene for bruk av stamceller er ganske klare, men de moralske og etiske aspektene ved dette problemet diskuteres fortsatt, siden det i de fleste tilfeller brukes embryonale stamceller hentet fra menneskelige embryoer drept under abort.
Varigheten av interfasen i plante- og dyreceller er i gjennomsnitt 10-20 timer, mens mitose tar ca. 1-2 timer.
I løpet av påfølgende delinger i flercellede organismer blir datterceller mer og mer mangfoldige, ettersom de leser informasjon fra et økende antall gener.
Noen celler slutter til slutt å dele seg og dør, noe som kan skyldes fullføringen av visse funksjoner, som i tilfelle av epidermale celler i huden og blodcellene, eller skade på disse cellene av miljøfaktorer, spesielt patogener. Genetisk programmert celledød kalles apoptose mens tilfeldig død er nekrose.
Mitose er delingen av somatiske celler. Faser av mitose
Mitose- en metode for indirekte deling av somatiske celler.
Under mitose går cellen gjennom en rekke påfølgende faser, som et resultat av at hver dattercelle mottar samme sett med kromosomer som i modercellen.
Mitose er delt inn i fire hovedfaser: profase, metafase, anafase og telofase. Profase- det lengste stadiet av mitose, hvor det oppstår kromatinkondensasjon, som et resultat av at X-formede kromosomer, bestående av to kromatider (datterkromosomer), blir synlige. I dette tilfellet forsvinner nukleolen, sentriolene divergerer mot cellens poler, og akromatinspindelen (spindelen) til mikrotubuli begynner å dannes. På slutten av profasen brytes kjernemembranen opp i separate vesikler.

PÅ metafase kromosomer står på linje langs ekvator av cellen med sine sentromerer, som mikrotubuli av en fullt dannet delingsspindel er festet til. På dette stadiet av delingen er kromosomene tettest og har en karakteristisk form, noe som gjør det mulig å studere karyotypen.
PÅ anafase rask DNA-replikasjon skjer i sentromerene, som et resultat av at kromosomene deler seg og kromatidene divergerer mot cellens poler, strukket av mikrotubuli. Fordelingen av kromatider må være helt lik, siden det er denne prosessen som opprettholder konstanten av antall kromosomer i cellene i kroppen.
På scenen telofase datterkromosomer samles ved polene, despiraliserer, rundt dem dannes det kjernekonvolutter fra vesiklene, og nukleoler vises i de nydannede kjernene.
Etter deling av kjernen skjer deling av cytoplasma - cytokinese, hvor det er en mer eller mindre jevn fordeling av alle organellene i modercellen.
Som et resultat av mitose dannes altså to datterceller fra én modercelle, som hver er en genetisk kopi av modercellen (2n2c).
I syke, skadede, aldrende celler og spesialiserte vev i kroppen kan det oppstå en litt annen delingsprosess - amitose. Amitose kalt direkte deling av eukaryote celler, der dannelsen av genetisk ekvivalente celler ikke skjer, siden cellekomponentene er ujevnt fordelt. Det forekommer i planter i endospermen og hos dyr i leveren, brusken og hornhinnen i øyet.
Meiose. Faser av meiose
Meiose- dette er en metode for indirekte deling av primære kjønnsceller (2n2c), som et resultat av at haploide celler (1n1c), oftest kjønnsceller, dannes.
I motsetning til mitose, består meiose av to påfølgende celledelinger, hver etterfulgt av en interfase. Den første deling av meiose (meiose I) kalles reduksjon, siden i dette tilfellet er antall kromosomer halvert, og den andre divisjonen (meiose II) - likning, siden antallet kromosomer er bevart i prosessen.

Interfase I fortsetter på samme måte som interfasen av mitose. Meiose I er delt inn i fire faser: profase I, metafase I, anafase I og telofase I. profeti I To hovedprosesser skjer: konjugering og kryssing. Konjugasjon- dette er prosessen med fusjon av homologe (parrede) kromosomer langs hele lengden. Kromosomparene dannet under konjugering beholdes til slutten av metafase I.
Krysser over- gjensidig utveksling av homologe regioner av homologe kromosomer. Som et resultat av kryssing får kromosomene som organismen mottar fra begge foreldrene nye kombinasjoner av gener, noe som fører til utseendet til genetisk mangfoldig avkom. På slutten av profase I, som i mitoseprofasen, forsvinner nukleolen, sentriolene divergerer mot cellens poler, og kjernehylsen går i oppløsning.

PÅ metafase I par av kromosomer stiller opp langs ekvator av cellen, mikrotubuli av fisjonsspindelen er festet til sentromerene deres.
PÅ anafase I hele homologe kromosomer bestående av to kromatider divergerer til polene.
PÅ telofase I rundt klynger av kromosomer ved cellens poler, dannes kjernemembraner, dannes nukleoler.
Cytokinese I gir deling av cytoplasma til datterceller.
Dattercellene dannet som et resultat av meiose I (1n2c) er genetisk heterogene, siden deres kromosomer, tilfeldig spredt til cellens poler, inneholder ulike gener.
Sammenlignende egenskaper ved mitose og meiose
| skilt | Mitose | Meiose | |
| Hvilke celler begynner å dele seg? | Somatisk (2n) | Primære kjønnsceller (2n) | |
| Antall divisjoner | 1 | 2 | |
| Hvor mange og hva slags celler dannes i delingsprosessen? | 2 somatiske (2n) | 4 seksuell (n) | |
| Interfase | Cellepreparat for deling, DNA-duplisering | Veldig kort, DNA-duplisering forekommer ikke | |
| Faser | Meiose I | Meiose II | |
| Profase | Kromosomkondensering, forsvinning av kjernen, desintegrering av kjernekappen, konjugering og kryssing kan forekomme | Kondensering av kromosomer, forsvinning av kjernen, desintegrering av kjernekonvolutten | |
| metafase | Par av kromosomer er plassert langs ekvator, en delingsspindel dannes | Kromosomer står på linje langs ekvator, delingsspindelen dannes | |
| Anafase | Homologe kromosomer fra to kromatider divergerer mot polene | Kromatider divergerer mot polene | |
| Telofase | Kromosomer despiraliserer, nye kjernekonvolutter og nukleoler dannes | Kromosomer despiraliserer, nye kjernekonvolutter og nukleoler dannes | |
Interfase II veldig kort, siden DNA-dobling ikke forekommer i den, det vil si at det ikke er noen S-periode.
Meiose II også delt inn i fire faser: profase II, metafase II, anafase II og telofase II. PÅ profeti II de samme prosessene skjer som i profase I, med unntak av konjugering og kryssing.
PÅ metafase II Kromosomer er plassert langs ekvator av cellen.
PÅ anafase II Kromosomene deler seg ved sentromeren og kromatidene strekker seg mot polene.
PÅ telofase II kjernemembraner og nukleoler dannes rundt klynger av datterkromosomer.
Etter cytokinese II den genetiske formelen til alle fire dattercellene er 1n1c, men de har alle et annet sett med gener, som er et resultat av kryssing og en tilfeldig kombinasjon av mors og fars kromosomer i datterceller.
Utviklingen av kjønnsceller i planter og dyr
Gametogenese(fra gresk. kjønnsceller- kone, kjønnsceller- mann og genesis- opprinnelse, forekomst) er prosessen med dannelse av modne kjønnsceller.
Siden seksuell reproduksjon oftest krever to individer - kvinnelige og mannlige, som produserer forskjellige kjønnsceller - egg og sædceller, bør prosessene for dannelse av disse gametene være forskjellige.
Prosessens karakter avhenger også i stor grad av om den skjer i en plante- eller dyrecelle, siden det i planter kun skjer mitose under dannelsen av kjønnsceller, mens det hos dyr forekommer både mitose og meiose.
Utviklingen av kjønnsceller i planter. Hos angiospermer skjer dannelsen av mannlige og kvinnelige kjønnsceller i forskjellige deler av blomsten - henholdsvis støvbærere og pistiller.
Før dannelsen av mannlige kjønnsceller - mikrogametogenese(fra gresk. mikros- liten) - skjer mikrosporogenese, det vil si dannelsen av mikrosporer i støvbærerens støvbærere. Denne prosessen er assosiert med den meiotiske deling av modercellen, som resulterer i fire haploide mikrosporer. Mikrogametogenese er assosiert med mitotisk deling av mikrosporer, noe som gir en mannlig gametofytt av to celler - en stor vegetativ(sifonogen) og grunne generativ. Etter deling er hanngametofytten dekket med tette skall og danner et pollenkorn. I noen tilfeller, selv i prosessen med pollenmodning, og noen ganger først etter overføring til pistillens stigma, deler den generative cellen seg mitotisk med dannelsen av to immobile mannlige kjønnsceller - sperm. Etter pollinering dannes et pollenrør fra den vegetative cellen, gjennom hvilken sæd trenger inn i eggstokken til pistillen for befruktning.

Utviklingen av kvinnelige kjønnsceller i planter kalles megagametogenese(fra gresk. megas- stor). Det forekommer i eggstokken til pistillen, som er innledet med megasporogenese, som et resultat av hvilket fire megasporer dannes fra modercellen til megasporen som ligger i kjernen ved meiotisk deling. En av megasporene deler seg mitotisk tre ganger, og gir opphav til den kvinnelige gametofytten, en embryosekk med åtte kjerner. Med den påfølgende isoleringen av cytoplasmaene til dattercellene, blir en av de resulterende cellene et egg, på sidene som ligger de såkalte synergidene, tre antipoder dannes i motsatt ende av embryoposen, og i midten , som et resultat av fusjonen av to haploide kjerner, dannes en diploid sentralcelle.

Utviklingen av kjønnsceller hos dyr. Hos dyr skilles to prosesser for dannelse av kjønnsceller - spermatogenese og oogenese.
spermatogenese(fra gresk. sperm, spermatos- frø og genesis- opprinnelse, forekomst) er prosessen med dannelse av modne mannlige kjønnsceller - spermatozoer. Hos mennesker forekommer det i testiklene, eller testiklene, og er delt inn i fire perioder: reproduksjon, vekst, modning og dannelse.

PÅ hekkesesongen primordiale kjønnsceller deler seg mitotisk, noe som resulterer i dannelsen av diploid spermatogoni. PÅ vekstperiode spermatogonia akkumulerer næringsstoffer i cytoplasmaet, øker i størrelse og blir til primære spermatocytter, eller spermatocytter av 1. orden. Først etter det går de inn i meiose ( modningsperiode), som først resulterer i to sekundær spermatocytt, eller spermatocytt av 2. orden, og deretter - fire haploide celler med en ganske stor mengde cytoplasma - spermatider. PÅ formasjonsperiode de mister nesten hele cytoplasmaet og danner et flagell som blir til spermatozoer.
sædceller, eller gummier,- veldig små mobile mannlige kjønnsceller med hode, nakke og hale.

PÅ hode, bortsett fra kjernen, er akrosom- et modifisert Golgi-kompleks, som sikrer oppløsning av eggets membraner under befruktning. PÅ nakke det er sentrioler av cellesenteret, og grunnlaget hestehale danner mikrotubuli som direkte støtter bevegelsen til sædcellene. Den inneholder også mitokondrier, som gir sædcellene ATP-energi for bevegelse.
Ovogenese(fra gresk. FN- et egg og genesis- opprinnelse, forekomst) er prosessen med dannelse av modne kvinnelige kjønnsceller - egg. Hos mennesker forekommer den i eggstokkene og består av tre perioder: reproduksjon, vekst og modning. Perioder med reproduksjon og vekst, lik de i spermatogenese, forekommer selv under intrauterin utvikling. Samtidig dannes diploide celler fra de primære kjønnscellene som følge av mitose. oogonia, som deretter blir til diploid primær oocytter, eller oocytter av 1. orden. Meiose og påfølgende cytokinese som forekommer i modningsperiode, er preget av ujevn deling av cytoplasmaet til modercellen, slik at som et resultat først oppnås en sekundær oocytt, eller oocytt 2. orden, og første polare legeme, og deretter fra den sekundære oocytten - egget, som beholder hele tilførselen av næringsstoffer, og den andre polare kroppen, mens den første polare kroppen er delt i to. Polare kropper tar bort overflødig genetisk materiale.
Hos mennesker produseres egg med et intervall på 28-29 dager. Syklusen knyttet til modning og frigjøring av egg kalles menstruasjonssyklusen.
Egg- en stor kvinnelig kjønnscelle, som ikke bare bærer et haploid sett med kromosomer, men også en betydelig tilførsel av næringsstoffer for den påfølgende utviklingen av embryoet.

Egget hos pattedyr er dekket med fire membraner, som reduserer sannsynligheten for skade på det av ulike faktorer. Diameteren på egget hos mennesker når 150-200 mikron, mens det hos en struts kan være flere centimeter.
Celledeling er grunnlaget for vekst, utvikling og reproduksjon av organismer. Rollen til mitose og meiose
Hvis celledeling i encellede organismer fører til en økning i antall individer, det vil si reproduksjon, kan denne prosessen i flercellede organismer ha en annen betydning. Således er celledeling av embryoet, med utgangspunkt i zygoten, det biologiske grunnlaget for de sammenkoblede prosessene for vekst og utvikling. Lignende endringer observeres hos en person i ungdomsårene, når antall celler ikke bare øker, men også en kvalitativ endring i kroppen oppstår. Reproduksjon av flercellede organismer er også basert på celledeling, for eksempel under aseksuell reproduksjon, på grunn av denne prosessen gjenopprettes en hel kropp fra en del av organismen, og under seksuell reproduksjon dannes kjønnsceller under gametogenese, som deretter gir en ny organisme. Det skal bemerkes at de viktigste metodene for eukaryotisk celledeling - mitose og meiose - har forskjellig betydning i livssyklusene til organismer.
Som et resultat av mitose er det en jevn fordeling av arvelig materiale mellom datterceller - eksakte kopier av moren. Uten mitose ville eksistensen og veksten av flercellede organismer som utvikler seg fra en enkelt celle, zygoten, vært umulig, siden alle celler i slike organismer må inneholde den samme genetiske informasjonen.
I prosessen med deling blir datterceller mer og mer forskjellige i struktur og funksjoner, noe som er assosiert med aktivering av nye grupper av gener i dem på grunn av intercellulær interaksjon. Dermed er mitose nødvendig for utviklingen av en organisme.
Denne metoden for celledeling er nødvendig for prosessene med aseksuell reproduksjon og regenerering (gjenoppretting) av skadet vev, så vel som organer.
Meiose sikrer på sin side konstansen til karyotypen under seksuell reproduksjon, da den reduserer med halvparten av settet med kromosomer før seksuell reproduksjon, som deretter gjenopprettes som et resultat av befruktning. I tillegg fører meiose til oppkomsten av nye kombinasjoner av foreldregener på grunn av kryssing og tilfeldig kombinasjon av kromosomer i datterceller. Takket være dette er avkommet genetisk mangfoldig, noe som gir materiale for naturlig utvalg og er det materielle grunnlaget for evolusjonen. En endring i antall, form og størrelse på kromosomer, på den ene siden, kan føre til utseendet av forskjellige avvik i utviklingen av organismen og til og med dens død, og på den annen side kan det føre til utseendet til individer mer tilpasset miljøet.
Dermed er cellen en enhet for vekst, utvikling og reproduksjon av organismer.