Intern struktur af prokaryoter. Sammenlignende karakteristika af cellestrukturen af prokaryoter og eukaryoter
Prokaryote celler- det er de mest primitive, meget simpelt strukturerede organismer, der bevarer træk fra den dybe antikke. TIL prokaryot(eller prænukleære) organismer omfatter bakterier og blågrønalger (cyanobakterier). Baseret på ligheden i struktur og skarpe forskelle fra andre celler, klassificeres prokaryoter i det uafhængige kongerige af knuste celler.
Lad os se på strukturen prokaryot celle bruge bakterier som eksempel. Det genetiske apparat af en prokaryot celle er repræsenteret af DNA'et af et enkelt cirkulært kromosom, er placeret i cytoplasmaet og er ikke afgrænset fra det af en membran. Denne analog af kernen kaldes en nukleoid. DNA danner ikke komplekser med proteiner og derfor "virker" alle gener, der er en del af kromosomet, dvs. information læses løbende fra dem.
Prokaryot celle omgivet af en membran, der adskiller cytoplasmaet fra cellevæggen, dannet af et komplekst, meget polymert stof. Der er få organeller i cytoplasmaet, men der er talrige små ribosomer til stede (bakterieceller indeholder fra 5.000 til 50.000 ribosomer).
Struktur af en prokaryot celle
Cytoplasmaet i en prokaryot celle gennemtrænges af membraner, der danner det endoplasmatiske retikulum; det indeholder ribosomer, der udfører proteinsyntese.
Det indre af cellevæggen i en prokaryot celle er repræsenteret ved plasma membran, hvis fremspring ind i cytoplasmaet danner mesosomer, som er involveret i konstruktionen af cellulære partitioner, reproduktion og er stedet for DNA-vedhæftning. Respiration hos bakterier forekommer i mesosomer og i blågrønalger i cytoplasmatiske membraner.
Mange bakterier afsætter reservestoffer inde i cellen: polysaccharider, fedtstoffer, polyphosphater. Reservestoffer, når de indgår i stofskiftet, kan forlænge en celles levetid i fravær af eksterne energikilder.
(1-cellevæg, 2-ydre cytoplasmatisk membran, 3-kromosom (cirkulært DNA-molekyle), 4-ribosom, 5-mesosom, 6-invagination af den ydre cytoplasmatiske membran, 7-vakuoler, 8-flagella, 9-stabler af membraner, hvori fotosyntesen finder sted)
Som regel formerer bakterier sig ved at dele sig i to. Efter celleforlængelse dannes der gradvist en tværgående skillevæg, som lægges i retning fra ydersiden til indersiden, derefter spredes dattercellerne eller forbliver forbundet i karakteristiske grupper - kæder, pakker osv. Bakterier - coli Hvert 20. minut fordobler den sine tal.
Bakterier er karakteriseret ved sporedannelse. Det begynder med løsrivelsen af en del af cytoplasmaet fra modercellen. Den løsrevne del indeholder ét genom og er omgivet af en cytoplasmatisk membran. Så vokser en cellevæg, ofte flerlaget, omkring sporen. Hos bakterier sker den seksuelle proces i form af en udveksling af genetisk information mellem to celler. Den seksuelle proces øger den arvelige variabilitet af mikroorganismer.
De fleste levende organismer er forenet i superriget af eukaryoter, som omfatter riget af planter, svampe og dyr. Eukaryote celler er større prokaryote celler, består af et overfladeapparat, en kerne og et cytoplasma.
Eukaryot celle
Eukaryot(eukaryot)celler indeholder en kerne koordinerer den vitale aktivitet af cellen, hvori kroppens arvelige apparat er placeret, og talrige organoider, udfører forskellige funktioner. De fleste eukaryoter er aerobe, det vil sige, at de bruger atmosfærisk ilt til energimetabolisme.
Den mest åbenlyse Forskellen mellem prokaryoter og eukaryoter er, at sidstnævnte har en kerne, hvilket afspejles i navnene på disse grupper: "karyo" er oversat fra oldgræsk som kerne, "pro" - før, "eu" - godt. Derfor er prokaryoter prænukleære organismer, eukaryoter er nukleare.
Dette er dog langt fra den eneste og måske ikke den vigtigste forskel mellem prokaryote organismer og eukaryoter. Prokaryote celler har slet ikke membranorganeller.(med sjældne undtagelser) - mitokondrier, kloroplaster, Golgi-kompleks, endoplasmatisk retikulum, lysosomer. Deres funktioner udføres af udvækster (invaginationer) af cellemembranen, hvorpå der er placeret forskellige pigmenter og enzymer, der sikrer vitale processer.
Prokaryoter har ikke de kromosomer, der er karakteristiske for eukaryoter. Deres vigtigste genetiske materiale er nukleoid, normalt formet som en ring. I eukaryote celler er kromosomer komplekser af DNA og histonproteiner (leg vigtig rolle i DNA-emballage). Disse kemiske komplekser kaldes kromatin. Nukleoiden af prokaryoter indeholder ikke histoner, og de RNA-molekyler, der er forbundet med den, giver den sin form.
Eukaryote kromosomer findes i kernen. I prokaryoter er nukleoiden placeret i cytoplasmaet og er normalt knyttet ét sted til cellemembranen.
Ud over nukleoiden har prokaryote celler forskellige mængder plasmider- nukleoider væsentligt mindre i størrelse end den vigtigste.
Antallet af gener i nukleoidet af prokaryoter er en størrelsesorden mindre end i kromosomer. Eukaryoter har mange gener, der udfører en regulerende funktion i forhold til andre gener. Dette gør det muligt for eukaryote celler af en flercellet organisme, der indeholder den samme genetiske information, at specialisere sig; ved at ændre dit stofskifte, reagere mere fleksibelt på ændringer i ydre og indre miljø. Strukturen af generne er også anderledes. I prokaryoter er gener i DNA arrangeret i grupper kaldet operoner. Hver operon transskriberes som en enkelt enhed.
Der er også forskelle mellem prokaryoter og eukaryoter i processerne for transkription og translation. Det vigtigste er, at i prokaryote celler kan disse processer forekomme samtidigt på ét molekyle af messenger (budbringer) RNA: mens det stadig syntetiseres på DNA, "sidder" ribosomer allerede i den færdige ende og syntetiserer protein. I eukaryote celler gennemgår mRNA såkaldt modning efter transkription. Og først efter det kan protein syntetiseres på det.
Ribosomerne af prokaryoter er mindre (sedimentationskoefficient 70S) end eukaryoters (80S). Antallet af proteiner og RNA-molekyler i de ribosomale underenheder er forskelligt. Det skal bemærkes, at ribosomerne (såvel som det genetiske materiale) i mitokondrier og kloroplaster ligner prokaryoter, hvilket kan indikere deres oprindelse fra gamle prokaryote organismer, der endte inde i værtscellen.
Prokaryoter er normalt kendetegnet ved en mere kompleks struktur af deres skaller. Ud over den cytoplasmatiske membran og cellevæggen har de også en kapsel og andre strukturer, afhængigt af typen af prokaryot organisme. Cellevæggen udfører en understøttende funktion og forhindrer penetrering skadelige stoffer. Bakteriecellevæggen indeholder murein (et glycopeptid). Blandt eukaryoter har planter en cellevæg (dens hovedkomponent er cellulose), og svampe har kitin.
Prokaryote celler deler sig ved binær fission. De har Ingen komplekse processer celledeling (mitose og meiose), karakteristisk for eukaryoter. Selvom nukleoidet før deling fordobles, ligesom kromatin i kromosomer. I livscyklus I eukaryoter er der en vekslen mellem diploide og haploide faser. I dette tilfælde dominerer den diploide fase normalt. I modsætning til dem har prokaryoter ikke dette.
Eukaryote celler varierer i størrelse, men under alle omstændigheder er de væsentligt større end prokaryote celler (tivis af gange).
Næringsstoffer kommer kun ind i prokaryote celler gennem osmose. I eukaryote celler kan der desuden også observeres fago- og pinocytose (“indfangningen” af mad og væske ved hjælp af den cytoplasmatiske membran).
Generelt ligger forskellen mellem prokaryoter og eukaryoter i sidstnævntes klart mere komplekse struktur. Det menes, at prokaryote celler er opstået gennem abiogenese (langsigtet kemisk udvikling under betingelserne i den tidlige Jord). Eukaryoter dukkede senere op fra prokaryoter gennem deres forening (symbiotiske og også kimære hypoteser) eller udviklingen af individuelle repræsentanter (invaginationshypotese). Kompleksiteten af eukaryote celler tillod dem at organisere en flercellet organisme og i evolutionsprocessen levere al den grundlæggende mangfoldighed af liv på Jorden.
Tabel over forskelle mellem prokaryoter og eukaryoter
| Skilt | Prokaryoter | Eukaryoter |
|---|---|---|
| Cellekerne | Ingen | Spise |
| Membranorganeller | Ingen. Deres funktioner udføres af invaginationer af cellemembranen, hvorpå pigmenter og enzymer er placeret. | Mitokondrier, plastider, lysosomer, ER, Golgi-kompleks |
| Cellemembraner | Mere kompleks er der forskellige kapsler. Cellevæggen er lavet af murein. | Hovedbestanddelen af cellevæggen er cellulose (i planter) eller kitin (i svampe). Dyreceller har ikke en cellevæg. |
| Genetisk materiale | Væsentlig mindre. Det er repræsenteret af en nukleoid og plasmider, som har en ringform og er placeret i cytoplasmaet. | Mængden af arvelig information er betydelig. Kromosomer (består af DNA og proteiner). Diploidi er karakteristisk. |
| Division | Binær celledeling. | Der er mitose og meiose. |
| Multicellularitet | Ikke typisk for prokaryoter. | De er repræsenteret af både encellede og flercellede former. |
| Ribosomer | Mindre | Større |
| Metabolisme | Mere forskelligartet (heterotrofer, fotosyntetisk og kemosyntetisk forskellige veje autotrofer; anaerob og aerob respiration). | Autotrofi forekommer kun i planter på grund af fotosyntese. Næsten alle eukaryoter er aerobe. |
| Oprindelse | Fra den livløse natur i processen med kemisk og præbiologisk udvikling. | Fra prokaryoter i gang med deres biologiske udvikling. |
Den prokaryote celle er meget enklere end dyre- og planteceller. På ydersiden er den dækket af en cellevæg, der udfører beskyttende, dannende og transporterende funktioner. Stivheden af cellevæggen tilvejebringes af murein. Nogle gange er bakteriecellen dækket ovenpå med en kapsel eller slimlag.
Bakteriens protoplasma er ligesom eukaryoter omgivet plasma membran. Sackulære, rørformede eller lamellære invaginationer af membranen indeholder mesosomer involveret i respirationsprocessen, bakteriochlorophyll og andre pigmenter. Det genetiske materiale af prokaryoter danner ikke en kerne, men er placeret direkte i cytoplasmaet. Bakterie-DNA er et enkelt cirkulært molekyle, som hver består af tusinder og millioner af nukleotidpar. Genomet af en bakteriecelle er meget enklere end cellerne fra mere udviklede væsner: i gennemsnit indeholder bakterielt DNA flere tusinde gener.
Fraværende i prokaryote celler endoplasmatisk retikulum, A ribosomer flyder frit i cytoplasmaet. Prokaryoter har ikke mitokondrier; Deres funktioner udføres delvist af cellemembranen.
Prokaryoter
Bakterier er den mindste af organismer med en cellulær struktur; deres størrelser spænder fra 0,1 til 10 mikron. Et typisk printsted kan rumme hundredtusindvis af mellemstore bakterier. Bakterier kan kun ses gennem et mikroskop, og derfor kaldes de mikroorganismer eller mikrober; mikroorganismer bliver undersøgt mikrobiologi . Den gren af mikrobiologi, der studerer bakterier, kaldes bakteriologi . Denne videnskab begyndte Anthony van Leeuwenhoek i det 17. århundrede.
Bakterie - de ældste kendte organismer. Spor af den vitale aktivitet af bakterier og blågrønalger (stromatolitter) tilhører arkæerne og dateres tilbage til 3,5 milliarder år gamle.På grund af muligheden for genudveksling mellem repræsentanter forskellige typer og endda slægter, er det ret svært at systematisere prokaryoter. En tilfredsstillende taksonomi af prokaryoter er endnu ikke blevet konstrueret; alle eksisterende systemer er kunstige og klassificerer bakterier i henhold til en gruppe af karakteristika uden at tage hensyn til deres fylogenetiske forhold. Tidligere bakterier sammen med svampe Og alger inkluderet i underriget af lavere planter. I øjeblikket er bakterier klassificeret som et separat superrige af prokaryoter. Det mest almindelige klassifikationssystem er Bergey system, som er baseret på cellevæggens struktur.
I slutningen af det 20. århundrede opdagede videnskabsmænd, at celler fra en relativt lidt undersøgt gruppe af bakterier - arkæbakterier – indeholde rRNA, forskellig i struktur fra både r-RNA af prokaryoter og r-RNA af eukaryoter. Strukturen af det genetiske apparat af arkæbakterier (tilstedeværelse introner og gentagne sekvenser, forarbejdning, form ribosomer) bringer dem tættere på eukaryoter; på den anden side har arkæbakterier også typiske træk ved prokaryoter (fravær af en kerne i cellen, tilstedeværelse af flageller, plasmider og gasvakuoler, rRNA-størrelse, nitrogenfiksering). Endelig adskiller arkæbakterier sig fra alle andre organismer i strukturen af deres cellevæg, typen af fotosyntese og nogle andre egenskaber. Arkæbakterier er i stand til at eksistere under ekstreme forhold (for eksempel i varme kilder ved temperaturer over 100 ° C, i havets dybder ved et tryk på 260 atm, i mættede saltopløsninger (30% NaCl)). Nogle arkæbakterier producerer metan, andre bruger svovlforbindelser til at producere energi.
Tilsyneladende er arkæbakterier en meget gammel gruppe af organismer; "ekstreme" muligheder angiver de forhold, der er karakteristiske for Jordens overflade i Arkæisk æra. Det menes, at arkæbakterier er tættest på de hypotetiske "pro-celler", der efterfølgende gav anledning til al mangfoldigheden af liv på Jorden.
For nylig er det blevet klart, at der er tre hovedtyper rRNA, præsenteret henholdsvis den første - i eukaryote celler, den anden - i cellerne af rigtige bakterier, såvel som i mitokondrier Og kloroplaster eukaryoter, den tredje - i arkæbakterier. Forskning i molekylær genetik har tvunget os til at tage et nyt blik på teorien om oprindelsen af eukaryoter. Det menes nu, at tre forskellige grene af prokaryoter udviklede sig samtidigt på oldtidens Jord - arkæbakterier, eubakterier og urkaryoter , kendetegnet ved forskellige strukturer og forskellige metoder til at opnå energi. Urkaryoter, som i det væsentlige var den nuklear-cytoplasmatiske komponent af eukaryoter, blev efterfølgende inkluderet som symbionter repræsentanter for forskellige grupper af eubakterier, som blev til mitokondrier og kloroplaster af fremtidige eukaryote celler.
Således er den klasserang, der tidligere blev tildelt for arkæbakterier, klart utilstrækkelig. I øjeblikket har mange forskere en tendens til at opdele prokaryoter i to riger: arkæbakterier og rigtige bakterier (eubakterier ) eller endda adskille archaebacteria i et separat superrige Archaea.
Klassificeringen af rigtige bakterier er givet i ordning.
I bakteriecelle Der er ingen kerne, kromosomerne er frit placeret i cytoplasmaet. Derudover mangler bakteriecellen membranorganeller: mitokondrier, EPS, Golgi apparat osv. Ydersiden af cellemembranen er dækket af en cellevæg.
De fleste bakterier bevæger sig passivt ved hjælp af vand eller luftstrømme. Kun nogle af dem har bevægelsesorganeller - flageller . Prokaryote flageller er meget enkle i strukturen og består af flagellinproteinet, som danner en hul cylinder med en diameter på 10-20 nm. De skruer ind i mediet og driver cellen fremad. Tilsyneladende er dette den eneste kendte struktur i naturen, der bruger hjulprincippet.
Baseret på deres form er bakterier opdelt i flere grupper:
cocci (har en rund form);
baciller (har en stavformet form);
spirilla (har form som en spiral);
vibrios (har form af et komma).
Baseret på respirationsmetoden opdeles bakterier i aerober (de fleste bakterier) og anaerobe (forårsager stivkrampe, botulisme, koldbrand). Førstnævnte har brug for ilt for at trække vejret; for sidstnævnte er ilt ubrugeligt eller endda giftigt.
Bakterier formerer sig ved at dele sig cirka hvert 20. minut (under gunstige forhold). DNA replikeres, hvor hver dattercelle modtager sin egen kopi af moder-DNA'et. Overførsel af DNA mellem ikke-delende celler er også mulig (gennem indfangning af "nøgent" DNA, vha. bakteriofager eller af konjugation , når bakterier er forbundet med hinanden af kopulatoriske fimbriae), sker der dog ikke en stigning i antallet af individer. Reproduktion forhindres af solens stråler og produkterne af deres egen vitale aktivitet.Bakteriers adfærd er ikke særlig kompleks. Kemiske receptorer registrerer ændringer i miljøets surhedsgrad og koncentrationen af forskellige stoffer: sukkerarter, aminosyrer, oxygen. Mange bakterier reagerer på ændringer i temperatur eller lys, og nogle bakterier kan mærke Jordens magnetfelt.
Under ugunstige forhold bliver bakterien dækket af en tæt skal, cytoplasmaet dehydreres, og vital aktivitet ophører næsten. I denne tilstand kan bakteriesporer forblive i et dybt vakuum i timevis og tåle temperaturer fra -240 °C til +100 °C.
1. En prokaryot celle er karakteriseret ved tilstedeværelsen
A) ribosomer
B) mitokondrier
B) dannet kerne
D) plasmamembran
D) endoplasmatisk retikulum
E) et cirkulært DNA
Svar
2. Prokaryote celler er forskellige fra eukaryote celler
A) tilstedeværelsen af ribosomer
B) fravær af mitokondrier
B) mangel på en dannet kerne
D) tilstedeværelsen af en plasmamembran
D) fravær af bevægelsesorganeller
E) tilstedeværelsen af et ringkromosom
Svar
3. Etabler en overensstemmelse mellem strukturen af celler og deres type: 1-prokaryot, 2-eukaryot
A) ikke har en dannet kerne
B) har en kernemembran
B) diploid eller haploid
D) altid haploid
D) har ikke mitokondrier eller Golgi-kompleks
E) indeholder mitokondrier, Golgi-kompleks
Svar
A1 B2 C2 D1 D1 E2
4. Hvorfor klassificeres bakterier som prokaryoter?
A) indeholder en kerne i cellen, adskilt fra cytoplasmaet
B) består af mange differentierede celler
B) har et ringkromosom
D) ikke har et cellecenter, Golgi-kompleks og mitokondrier
D) ikke har en kerne isoleret fra cytoplasmaet
E) har cytoplasma og plasmamembran
Svar
5. En bakteriecelle tilhører gruppen af prokaryote celler, da den
A) har ikke en skal-dækket kerne
B) har cytoplasma
B) har et DNA-molekyle nedsænket i cytoplasmaet
D) har en ydre plasmamembran
D) har ikke mitokondrier
E) har ribosomer, hvor proteinbiosyntese finder sted
Svar
6. Celler af eukaryote organismer, i modsætning til prokaryote, har
A) cytoplasma
B) kerne dækket med en skal
B) DNA-molekyler
D) mitokondrier
D) tæt skal
E) endoplasmatisk retikulum
Svar
7. Etabler en overensstemmelse mellem cellens karakteristika og dens type: 1-prokaryot, 2-eukaryot
A) Der er ingen membranorganeller
B) Der er en cellevæg lavet af murein
B) Arvemateriale er repræsenteret af en nukleoid
D) Indeholder kun små ribosomer
D) Arvemateriale er repræsenteret ved lineært DNA
E) Cellulær respiration forekommer i mitokondrier
Svar
A1 B1 C1 D1 D2 E2
8. Prokaryote celler er forskellige fra eukaryote celler
A) tilstedeværelsen af en nukleoid i cytoplasmaet
B) tilstedeværelsen af ribosomer i cytoplasmaet
B) ATP-syntese i mitokondrier
D) tilstedeværelsen af det endoplasmatiske retikulum
D) fravær af en morfologisk distinkt kerne
E) tilstedeværelsen af invaginationer af plasmamembranen, der udfører funktionen af membranorganeller
Ud fra deres cellestruktur opdeles levende organismer i prokaryot Og eukaryoter. Cellerne i begge er omgivet plasma membran, uden for hvilket der i mange tilfælde er cellevæg. Inde i cellen er der en halvvæske cytoplasma. Imidlertid er prokaryote celler meget enklere end eukaryote celler.
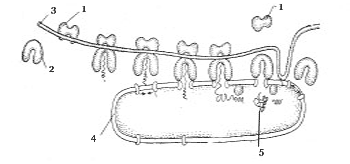
Grundlæggende genetisk materiale prokaryot (fra græsk om– før og karyon- kerne) er placeret i cytoplasmaet i form af et cirkulært DNA-molekyle. Dette molekyle ( nukleoid) er ikke omgivet af den nukleare kappe, der er karakteristisk for eukaryoter og er knyttet til plasmamembranen (fig. 1). Prokaryoter har således ikke en dannet kerne. Ud over nukleoiden kaldes et lille cirkulært DNA-molekyle plasmid. Plasmider kan bevæge sig fra en celle til en anden og integreres i hoved-DNA-molekylet.
Nogle prokaryoter har plasmamembranforlængelser: mesosomer, lamellære thylakoider, kromatoforer. De indeholder enzymer involveret i fotosyntese og respirationsprocesser. Derudover er mesosomer forbundet med DNA-syntese og proteinsekretion.
Prokaryote celler er små i størrelse, deres diameter er 0,3-5 μm. På ydersiden af plasmamembranen af alle prokaryoter (med undtagelse af mycoplasmas) er cellevæg. Den består af komplekser af proteiner og oligosaccharider, arrangeret i lag, beskytter cellen og bevarer sin form. Det er adskilt fra plasmamembranen af et lille intermembranrum.
Kun ikke-membranorganeller findes i cytoplasmaet af prokaryoter ribosomer. Strukturen af ribosomer af prokaryoter og eukaryoter er ens, dog er ribosomer af prokaryoter mindre og er ikke knyttet til membranen, men er placeret direkte i cytoplasmaet.

Mange prokaryoter er mobile og kan svømme eller glide ved hjælp af flageller.
Prokaryoter formerer sig normalt ved at dele sig i to ( binær). Forud for deling sker der et meget kort stadie af kromosomduplikation eller replikation. Så prokaryoter er haploide organismer.
Prokaryoter omfatter bakterier og blågrønalger eller cyanobakterier. Prokaryoter dukkede op på Jorden for omkring 3,5 milliarder år siden og var sandsynligvis de første cellulære livsformer, der gav anledning til moderne prokaryoter og eukaryoter.
Eukaryoter (fra græsk æj- rigtigt, karyon- kerne) i modsætning til prokaryoter, har en dannet kerne omgivet atomkappe– dobbeltlags membran. DNA-molekyler fundet i kernen er åbne (lineære molekyler). Ud over kernen er en del af den genetiske information indeholdt i mitokondriers og kloroplasters DNA. Eukaryoter dukkede op på Jorden for cirka 1,5 milliarder år siden.
I modsætning til prokaryoter, som er repræsenteret af solitære organismer og koloniale former, kan eukaryoter være encellede (for eksempel amøbe), koloniale (Volvox) og flercellede organismer. De er opdelt i tre store riger: Dyr, Planter og Svampe.
Diameteren af eukaryote celler er 5-80 μm. Ligesom prokaryote celler er eukaryote celler omgivet af plasma membran bestående af proteiner og lipider. Denne membran fungerer som en selektiv barriere, permeabel for nogle forbindelser og uigennemtrængelig for andre. Uden for plasmamembranen er en stærk cellevæg, som i planter hovedsageligt består af cellulosefibre, og i svampe - af kitin. Cellevæggens hovedfunktion er at sikre en konstant celleform. Da plasmamembranen er permeabel for vand, og plante- og svampeceller typisk kommer i kontakt med opløsninger med lavere ionstyrke end ionstyrken af opløsningen inde i cellen, vil vand strømme ind i cellerne. På grund af dette vil volumenet af celler stige, plasmamembranen vil begynde at strække sig og kan briste. Cellevæggen forhindrer celleudvidelse og ødelæggelse.
Dyr mangler en cellevæg, men det ydre lag af plasmamembranen er beriget med kulhydratkomponenter. Dette ydre lag af plasmamembranen af dyreceller kaldes glykokalyx. Celler fra flercellede dyr kræver ikke en stærk cellevæg, da der er andre mekanismer, der giver regulering af cellevolumen. Da cellerne i flercellede dyr og encellede organismer, der lever i havet, er i et miljø, hvor den samlede koncentration af ioner er tæt på den intracellulære ionkoncentration, cellerne svulmer ikke eller brister. Encellede dyr, der lever i ferskvand (amoebe, slipper ciliater) har kontraktile vakuoler, der konstant fjerner vandet, der kommer ind i cellen til ydersiden.
Strukturelle komponenter i en eukaryot celle
Inde i cellen, under plasmamembranen, er der cytoplasma. Hovedstoffet i cytoplasmaet (hyaloplasma) er en koncentreret opløsning af uorganiske og organiske forbindelser, hvis hovedkomponenter er proteiner. Det her kolloide system, som kan skifte fra flydende til geltilstand og tilbage. En væsentlig del af de cytoplasmatiske proteiner er enzymer, der udfører forskellige kemiske reaktioner. Beliggende i hyaloplasmaet organoider, udfører forskellige funktioner i cellen. Organeller kan være membran (kerne, Golgi-apparat, endoplasmatisk retikulum, lysosomer, mitokondrier, kloroplaster) og ikke-membran (cellulært center, ribosomer, cytoskelet).
Membranorganellerhovedbestanddelen af membranorganeller er membran. Biologiske membraner bygges iflg generelt princip, Men kemisk sammensætning membraner af forskellige organeller er forskellige. Alle cellemembraner er tynde film (7-10 nm tykke), hvis basis er et dobbeltlag af lipider (dobbeltlag), arrangeret således, at de ladede hydrofile dele af molekylerne er i kontakt med mediet og de hydrofobe rester fedtsyrer af hvert monolag er rettet ind i membranen og er i kontakt med hinanden (fig. 3). Proteinmolekyler (integrerede membranproteiner) er indbygget i lipid-dobbeltlaget på en sådan måde, at de hydrofobe dele af proteinmolekylet er i kontakt med fedtsyreresterne i lipidmolekylerne, og de hydrofile dele udsættes for miljøet. Derudover forbindes en del af de opløselige (ikke-membranproteiner) til membranen hovedsageligt på grund af ioniske interaktioner (perifere membranproteiner). Kulhydratfragmenter er også knyttet til mange proteiner og lipider i membraner. Dermed, biologiske membraner- Det er lipidfilm, hvori integrerede proteiner er indlejret.
En af membranernes hovedfunktioner er at skabe en grænse mellem cellen og miljøet og cellens forskellige rum. Lipid-dobbeltlaget er hovedsageligt permeabelt for fedtopløselige forbindelser og gasser; hydrofile stoffer transporteres gennem membraner ved hjælp af specielle mekanismer: lav molekylvægt - gennem forskellige bærere (kanaler, pumper osv.) og høj molekylvægt - gennem processer exo- Og endocytose(Fig. 4).

Ris. 4. Skema for stofoverførsel gennem membranen
På endocytose visse stoffer sorberes på membranens overflade (på grund af interaktion med membranproteiner). På dette tidspunkt dannes en invagination af membranen ind i cytoplasmaet. Et hætteglas indeholdende den overførte forbindelse adskilles derefter fra membranen. Dermed, endocytose– er overførsel af højmolekylære forbindelser ind i cellen ydre miljø, omgivet af en sektion af membran. Omvendt proces, det er eksocytose- Dette er overførsel af stoffer fra cellen til det ydre. Det sker ved fusion med plasmamembranen af en vesikel fyldt med transporterede stoffer. højmolekylære forbindelser. Vesiklens membran smelter sammen med plasmamembranen, og dens indhold hælder ud.
Kanaler, pumper og andre transportører er molekyler af integrerede membranproteiner, der typisk danner en pore i membranen.
Ud over funktionerne med at adskille rummet og sikre selektiv permeabilitet, er membraner i stand til at registrere signaler. Denne funktion udføres af receptorproteiner, der binder signalmolekyler. Individuelle membranproteiner er enzymer, der udfører specifikke kemiske reaktioner.
Kerne - en storcellet organel, omgivet af en nuklear kappe og sædvanligvis med en sfærisk form. Der er kun én kerne i en celle, og selvom der er multinukleære celler (celler i skeletmuskler, nogle svampe) eller dem uden en kerne (erythrocytter og blodplader fra pattedyr), opstår disse celler fra mononukleære precursorceller.
Kernens hovedfunktion er opbevaring, overførsel og salg af genetisk information. Her duplikeres DNA-molekyler, hvilket resulterer i, at datterceller ved deling modtager det samme genetiske materiale. I kernen, ved hjælp af individuelle sektioner af DNA-molekyler (gener) som en matrix, sker syntesen af RNA-molekyler: informations- (mRNA), transport (tRNA) og ribosomalt (rRNA), der er nødvendige for proteinsyntese. I kernen samles ribosomale underenheder af rRNA-molekyler og proteiner, som syntetiseres i cytoplasmaet og overføres til kernen.
Kernen består af kernekappen, kromatin (kromosomer), nukleolus og nukleoplasma (karyoplasma).

Ris. 5. Kromatinstruktur: 1 – nukleosom, 2 – DNA
Under et mikroskop er zoner med tæt stof synlige inde i kernen - kromatin. I ikke-delende celler fylder det ensartet volumen af kernen eller kondenserer visse steder i form af tættere områder og er godt farvet med basiske farvestoffer. Kromatin er et kompleks af DNA og proteiner (fig. 5), for det meste positivt ladet histoner.
Antallet af DNA-molekyler i kernen er lig med antallet af kromosomer. Antallet og formen af kromosomer er et unikt kendetegn for en art. Hvert kromosom indeholder et DNA-molekyle, bestående af to indbyrdes forbundne strenge og formet som en dobbelt helix 2 nm tyk. Dens længde overstiger væsentligt cellens diameter: den kan nå flere centimeter. DNA-molekylet er negativt ladet, så det kan først folde (kondensere) efter binding til positivt ladede histonproteiner (fig. 6).
For det første snoer en dobbeltstreng af DNA sig omkring individuelle histonblokke, som hver indeholder 8 proteinmolekyler, og danner en "perler på en streng"-struktur omkring 10 nm tyk. Perlerne kaldes nukleosomer. Som et resultat af dannelsen af nukleosomer falder længden af DNA-molekylet med cirka 7 gange. Dernæst folder tråden med nukleosomer sig og danner en reb-lignende struktur omkring 30 nm tyk. Dette reb, der er bøjet til løkker, er derefter knyttet til de proteiner, der danner grundlaget for kromosomet. Som et resultat dannes en struktur med en tykkelse på omkring 300 nm. Yderligere kondensering af denne struktur fører til dannelsen af et kromosom.
I perioden mellem delinger folder kromosomet sig delvist ud. Som et resultat af dette bliver enkelte sektioner af DNA-molekylet, der skal udtrykkes i en given celle, befriet for proteiner og strækket, hvilket gør det muligt at aflæse information fra dem gennem syntese af RNA-molekyler.
Nukleolus er en type skabelon-DNA, der er ansvarlig for rRNA-syntese og opsamlet i separate områder af kernen. Nucleolus er den tætteste struktur af kernen; det er ikke en separat organel, men repræsenterer et af kromosomets loci. Det producerer rRNA, som derefter danner et kompleks med proteiner, der danner ribosomale underenheder, der går ind i cytoplasmaet.
Ikke-histone nukleare proteiner danner et strukturelt netværk i kernen. Det er repræsenteret af et lag af fibriller, der ligger under den nukleare kappe. Et intranukleært fibrilnetværk er knyttet til det, hvortil kromatinfibriller er knyttet.
Det nukleare hylster består af to membraner: ydre og indre, adskilt af et intermembranrum. Den ydre membran er i kontakt med cytoplasmaet, den kan indeholde polyribosomer, og den kan selv passere ind i membranerne i det endoplasmatiske retikulum. Den indre membran er forbundet med kromatin. Kernekappen sikrer således fiksering af kromosomalt materiale i kernens tredimensionelle rum.
Kerneskallen har runde huller - nukleare porer(Fig. 7). I poreområdet lukker de ydre og indre membraner sig og danner huller fyldt med fibriller og granulat. Inde i poren er der et komplekst system af proteiner, der giver selektiv binding og transport af makromolekyler. Antallet af nukleare porer afhænger af intensiteten af cellemetabolisme.
Endoplasmatisk retikulum, eller endoplasmatisk retikulum (ER), er et bizart netværk af kanaler, vakuoler, fladtrykte sække, indbyrdes forbundet og adskilt fra hyaloplasmaet af en membran (fig. 8).
Skelne ru Og glat EPJ . På membranerne af den ru ER er der ribosomer(Fig. 9), som syntetiserer proteiner udskilt fra cellen eller integreret i plasmamembranen. Det nysyntetiserede protein forlader ribosomet og passerer gennem en speciel kanal ind i hulrummet endoplasmatisk retikulum, hvor det gennemgår post-translationel modifikation, for eksempel binding til kulhydrater, proteolytisk spaltning af en del af polypeptidkæden og dannelsen af S-S-bindinger mellem cysteinrester i kæden. Disse proteiner transporteres derefter til Golgi-komplekset, hvor de enten er en del af lysosomer eller sekretoriske granula. I begge tilfælde ender disse proteiner inde i en membranvesikel (vesikel).
Ris. 9. Skema for proteinsyntese i groft ER: 1 – lille og
2 - stor ribosomal underenhed; 3 - rRNA-molekyle;
4 – grov EPJ; 5 – nysyntetiseret protein
Den glatte ER mangler ribosomer. Dens hovedfunktion er lipidsyntese og kulhydratmetabolisme. Det er veludviklet, for eksempel i cellerne i binyrebarken, som indeholder enzymer, der sikrer syntesen af steroidhormoner. I den glatte ER i leverceller er der enzymer, der udfører oxidation (afgiftning) af hydrofobe forbindelser, der er fremmede for kroppen, såsom lægemidler.

Ris. 10. Golgi-apparat: 1 – vesikler; 2 – tanke
Golgi kompleks (Fig. 10) består af 5-10 flade membranafgrænsede hulrum placeret parallelt. Endedelene af disse skiveformede strukturer har forlængelser. Der kan være flere sådanne formationer i en celle. I området af Golgi-komplekset er der et stort antal af membranvesikler. Nogle af dem er løsrevet fra de terminale dele af hovedstrukturen i form af sekretoriske granuler og lysosomer. Nogle af de små vesikler (vesikler), der bærer proteiner syntetiseret i det ru ER, flytter til Golgi-komplekset og smelter sammen med det. Golgi-komplekset er således involveret i akkumulering og yderligere modifikation af produkter syntetiseret i den rå ER og deres sortering.

Ris. 11. Dannelse og funktioner af lysosomer: 1 – fagosom; 2 - pinocytotisk vesikel; 3 - primært lysosom; 4 – Golgi-apparater; 5 – sekundært lysosom
Lysosomer - disse er vakuoler (fig. 11), begrænset af en membran, som knopper fra Golgi-komplekset. Der er lysosomer nok indeni surt miljø(pH 4,9-5,2). Der er hydrolytiske enzymer, der nedbryder forskellige polymerer ved sur pH (proteaser, nukleaser, glucosidaser, fosfataser, lipaser). Disse primære lysosomer fusionerer med endocytiske vakuoler, der indeholder komponenter, der skal nedbrydes. Stoffer, der kommer ind i det sekundære lysosom, nedbrydes til monomerer og overføres gennem lysosommembranen til hyaloplasmaet. Lysosomer deltager således i processerne med intracellulær fordøjelse.
Mitokondrier omgivet af to membraner: den ydre, der adskiller mitokondriet fra hyaloplasmaet, og den indre, der afgrænser dets indre indhold. Mellem dem er der et intermembranrum 10-20 nm bredt. Den indre membran danner adskillige fremspring ( cristas). Denne membran indeholder enzymer, der sikrer oxidation af aminosyrer, sukkerarter, glycerol og fedtsyrer dannet uden for mitokondrierne (Krebs cyklus) og udfører elektronoverførsel i åndedrætskæden (diagram). På grund af overførsel af elektroner langs åndedrætskæden fra et højt til et lavere energiniveau, lagres en del af den frigivne frie energi i form af ATP, cellens universelle energivaluta. Således er mitokondriers hovedfunktion oxidation af forskellige substrater og syntese af ATP-molekyler.
Skema med to elektronoverførsel langs åndedrætskæden
Inde i mitokondriet er der et cirkulært DNA-molekyle, der koder for en del af mitokondrieproteinerne. I mitokondriers (matrix) indre rum er der ribosomer svarende til ribosomer af prokaryoter, som sikrer syntesen af disse proteiner.
Det faktum, at mitokondrier har deres eget cirkulære DNA og prokaryote ribosomer, har ført til den hypotese, at mitokondriet er en efterkommer af en ældgammel prokaryot celle, der engang kom ind i en eukaryotisk celle, og som i evolutionsprocessen påtog sig visse funktioner.
Ris. 12. Kloroplaster (A) og thylakoidmembraner (B)
Plastider – organeller plantecelle, som indeholder pigmenter. I kloroplaster indeholder klorofyl og carotenoider, kromoplaster– carotenoider, i leukoplaster ingen pigmenter. Plastider er omgivet af en dobbelt membran. Inde i dem er der et system af membraner i form af flade bobler kaldet thylakoider(Fig. 12). Thylakoiderne er arrangeret i stakke, der ligner stakke af plader. Pigmenterne er indlejret i thylakoidmembranerne. Deres hovedfunktion er absorptionen af lys, hvis energi ved hjælp af enzymer indbygget i thylakoidmembranen omdannes til en gradient af H + ioner på thylakoidmembranen. Ligesom mitokondrier har plastider deres eget cirkulære DNA og ribosomer af prokaryot-typen. Tilsyneladende er plastider også prokaryote organismer, der lever i symbiose med eukaryote celler.
Ribosomer er ikke-membranøse cellulære organeller, der findes i både pro- og eukaryote celler. Eukaryote ribosomer er større i størrelse end prokaryote, deres størrelse er 25x20x20 nm. Ribosomet består af store og små underenheder, der støder op til hinanden. Mellem underenhederne i et fungerende ribosom er der en streng af mRNA.
Hver ribosomale underenhed er lavet af rRNA, tæt pakket og forbundet med proteiner. Ribosomer kan være placeret frit i cytoplasmaet eller være forbundet med ER-membraner. Frie ribosomer kan være enkeltstående, men de kan danne polysomer, når flere ribosomer er placeret sekventielt på en streng af mRNA. Ribosomernes hovedfunktion er proteinsyntese.
Cytoskelet - Det her muskuloskeletale system celler, herunder protein filamentøse (fibrillære) formationer, som er rammen af cellen og udfører motorisk funktion. Cytoskeletstrukturer er dynamiske, de opstår og desintegrerer. Cytoskelettet er repræsenteret af tre typer formationer: mellemfilamenter(tråde med en diameter på 10 nm), mikrofilament s (tråde med en diameter på 5-7 nm) og mikrotubuli. Mellemfilamenter er ikke-forgrenede proteinstrukturer i form af tråde, ofte arrangeret i bundter. Deres proteinsammensætning varierer i forskellige væv: i epitelet består de af keratin, i fibroblaster - af vimentin, i muskelceller- fra desmin. Mellemfilamenter vil udføre en understøttende rammefunktion.
Mikrofilamenter - disse er fibrillære strukturer placeret direkte under plasmamembranen i form af bundter eller lag. De er tydeligt synlige i amøbens pseudopoder, i fibroblasternes bevægelsesprocesser og i tarmepitelets mikrovilli (fig. 13). Mikrofilamenter er bygget af de kontraktile proteiner actin og myosin og er det intracellulære kontraktile apparat.
Mikrotubuli er en del af både midlertidige og permanente cellestrukturer. Midlertidige omfatter delingsspindelen, elementer i cytoskelettet af celler mellem delinger, og permanente omfatter cilia, flageller og centrioler i cellecentret. Mikrotubuli er lige, hule cylindre med en diameter på omkring 24 nm; deres vægge er dannet af runde molekyler af proteinet tubulin. Under et elektronmikroskop kan det ses, at mikrotubuli-tværsnittet er dannet af 13 underenheder forbundet i en ring. Mikrotubuli er til stede i hyaloplasmaet af alle eukaryote celler. En af mikrotubuli's funktioner er at skabe en ramme inde i celler. Derudover bevæger små vesikler sig langs mikrotubuli, som på skinner.
Cellecenter består af to centrioler placeret vinkelret på hinanden og tilhørende mikrotubuli. Disse organeller i delende celler deltager i dannelsen af delingsspindelen. Grundlaget for centriolen er 9 tripletter af mikrotubuli placeret rundt om omkredsen, der danner en hul cylinder, 0,2 µm bred og 0,3-0,5 µm lang. Når celler forbereder sig på at dele sig, divergerer centriolerne og fordobles. Før mitose deltager centrioler i dannelsen af spindelmikrotubuli. Celler af højere planter har ikke centrioler, men de har et lignende organiseringscenter for mikrotubuli.