Intern struktur av prokaryoter. Sammenlignende egenskaper av strukturen til prokaryote og eukaryote celler
prokaryote celler- dette er de mest primitive, veldig enkelt arrangert, som bevarer funksjonene i antikken. Til prokaryot(eller pre-nukleære) organismer inkluderer bakterier og blågrønne alger (cyanobakterier). Basert på strukturens fellestrekk og skarpe forskjeller fra andre celler, blir prokaryote celler isolert til et uavhengig kongerike av hagler.
Vurder strukturen prokaryot celle på eksemplet med bakterier. Det genetiske apparatet til en prokaryot celle er representert av DNAet til et enkelt ringkromosom, lokalisert i cytoplasmaet og ikke avgrenset fra det av en membran. En slik analog av kjernen kalles en nukleoid. DNA danner ikke komplekser med proteiner, og derfor "fungerer" alle genene som utgjør kromosomet, d.v.s. informasjon leses fortløpende fra dem.
prokaryot celle omgitt av en membran som skiller cytoplasmaet fra celleveggen, dannet av et komplekst, høypolymert stoff. Det er få organeller i cytoplasmaet, men mange små ribosomer er tilstede (bakterieceller inneholder fra 5 000 til 50 000 ribosomer).
Strukturen til en prokaryot celle
Cytoplasmaet til en prokaryot celle er gjennomsyret av membraner som danner det endoplasmatiske retikulum, og det inneholder ribosomer som utfører proteinsyntese.
Den indre delen av celleveggen til en prokaryot celle er representert ved plasmamembran, fremspring som inn i cytoplasmaet danner mesosomer involvert i konstruksjonen av cellevegger, reproduksjon, og er stedet for DNA-feste. Respirasjon hos bakterier utføres i mesosomer, i blågrønne alger i cytoplasmatiske membraner.
Hos mange bakterier avsettes reservestoffer inne i cellen: polysakkarider, fett, polyfosfater. Reservestoffer, som er inkludert i metabolismen, kan forlenge cellens levetid i fravær av eksterne energikilder.
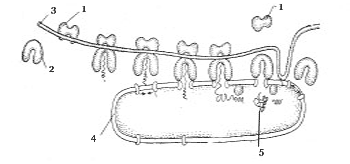
(1-cellevegg, 2-ytre cytoplasmatisk membran, 3-kromosom (sirkulært DNA-molekyl), 4-ribosom, 5-mesosom, 6-invaginasjon av den ytre cytoplasmatiske membranen, 7-vakuoler, 8-flagella, 9-stabler av membraner, der fotosyntesen finner sted)
Vanligvis formerer bakterier seg ved å dele seg i to. Etter forlengelse av cellen dannes gradvis et tverrgående skillevegg, som legges i retning fra utsiden til innsiden, deretter divergerer dattercellene eller forblir forbundet i karakteristiske grupper - kjeder, pakker, etc. bakterie - coli dobler innbyggertallet hvert 20. minutt.
Bakterier er preget av sporulering. Det begynner med at en del av cytoplasmaet løsner fra modercellen. Den løsrevne delen inneholder ett genom og er omgitt av en cytoplasmatisk membran. Da vokser en cellevegg, ofte flerlags, rundt sporen. Hos bakterier observeres den seksuelle prosessen i form av utveksling av genetisk informasjon mellom to celler. Den seksuelle prosessen øker den arvelige variasjonen til mikroorganismer.
De fleste levende organismer er forent i eukaryotes rike, inkludert riket av planter, sopp og dyr. eukaryote celler er større prokaryote celler, består av et overflateapparat, kjerne og cytoplasma.
eukaryot celle
eukaryote(eukaryot)celler inneholder en kjerne koordinere den vitale aktiviteten til cellen, der kroppens arvelige apparat er lokalisert, og mange organeller utføre ulike funksjoner. De fleste eukaryoter er aerobe, det vil si at de bruker atmosfærisk oksygen for energimetabolisme.
Den mest åpenbare Forskjellen mellom prokaryoter og eukaryoter er at sistnevnte har en kjerne, som gjenspeiles i navnet på disse gruppene: "karyo" er oversatt fra gammelgresk som kjernen, "pro" - før, "eu" - bra. Derfor er prokaryoter pre-nukleære organismer, eukaryoter er nukleære.
Dette er imidlertid langt fra den eneste og kanskje ikke hovedforskjellen mellom prokaryote organismer og eukaryoter. Det er ingen membranorganeller i prokaryote celler i det hele tatt.(med sjeldne unntak) - mitokondrier, kloroplaster, Golgi-kompleks, endoplasmatisk retikulum, lysosomer. Deres funksjoner utføres av utvekster (invaginasjoner) av cellemembranen, på hvilke ulike pigmenter og enzymer er plassert som gir vitale prosesser.
Prokaryoter har ikke eukaryote kromosomer. Deres viktigste genetiske materiale er nukleoid vanligvis i form av en ring. I eukaryote celler er kromosomer komplekser av DNA og histonproteiner (lek viktig rolle i DNA-emballasje). Disse kjemiske kompleksene kalles kromatin. Nukleoiden til prokaryoter inneholder ikke histoner, og RNA-molekylene knyttet til den gir den sin form.
Eukaryote kromosomer er lokalisert i kjernen. Hos prokaryoter er nukleoidet lokalisert i cytoplasmaet og er vanligvis festet på ett sted til cellemembranen.
I tillegg til nukleoiden har prokaryote celler en annen mengde plasmid- nukleoider av en betydelig mindre størrelse enn den viktigste.
Antall gener i nukleoiden til prokaryoter er en størrelsesorden mindre enn i kromosomene. Eukaryoter har mange gener som utfører en regulerende funksjon i forhold til andre gener. Dette gjør det mulig for eukaryote celler fra en flercellet organisme, som inneholder den samme genetiske informasjonen, å spesialisere seg; endre stoffskiftet, reagere mer fleksibelt på endringer i ytre og Internt miljø. Strukturen til genene er også annerledes. Hos prokaryoter er gener i DNA ordnet i grupper - operoner. Hver operon er transkribert som en enkelt enhet.
Det er også forskjeller mellom prokaryoter og eukaryoter i prosessene for transkripsjon og translasjon. Det viktigste er at i prokaryote celler kan disse prosessene foregå samtidig på ett molekyl av matrise (informasjons-) RNA: mens det fortsatt syntetiseres på DNA, "sitter" ribosomer allerede i den ferdige enden og syntetiserer protein. I eukaryote celler gjennomgår mRNA såkalt modning etter transkripsjon. Og først etter det kan protein syntetiseres på den.
Ribosomene til prokaryoter er mindre (sedimentasjonskoeffisient 70S) enn de til eukaryoter (80S). Antallet proteiner og RNA-molekyler i sammensetningen av ribosomunderenheter er forskjellig. Det skal bemerkes at ribosomene (så vel som det genetiske materialet) til mitokondrier og kloroplaster ligner på prokaryoter, noe som kan indikere deres opprinnelse fra eldgamle prokaryote organismer som var inne i vertscellen.
Prokaryoter er vanligvis forskjellige i den mer komplekse strukturen til skallene deres. I tillegg til den cytoplasmatiske membranen og celleveggen har de også en kapsel og andre formasjoner, avhengig av typen prokaryot organisme. Celleveggen fungerer som en støtte og hindrer penetrering skadelige stoffer. Bakteriecelleveggen inneholder murein (et glykopeptid). Blant eukaryoter har planter en cellevegg (hovedkomponenten er cellulose), sopp har kitin.
Prokaryote celler deler seg ved binær fisjon. De har Nei komplekse prosesser celledeling (mitose og meiose) karakteristisk for eukaryoter. Selv før deling dobles nukleoidet, akkurat som kromatin i kromosomer. PÅ Livssyklus eukaryoter har vekslende diploide og haploide faser. I dette tilfellet dominerer vanligvis den diploide fasen. I motsetning til dem har ikke prokaryoter dette.
Eukaryote celler varierer i størrelse, men i alle fall er de betydelig større enn prokaryote celler (ti titalls ganger).
Næringsstoffer De kommer inn i prokaryote celler bare ved osmose. I eukaryote celler kan i tillegg fago- og pinocytose ("fanging" av mat og væske ved hjelp av cytoplasmatisk membran) også observeres.
Generelt ligger forskjellen mellom prokaryoter og eukaryoter i den klart mer komplekse strukturen til sistnevnte. Det antas at celler av den prokaryote typen oppsto gjennom abiogenese (langsiktig kjemisk utvikling under forholdene til den tidlige jorden). Eukaryoter dukket senere opp fra prokaryoter, ved å kombinere dem (symbiotiske, så vel som kimære hypoteser) eller ved utviklingen av individuelle representanter (invaginasjonshypotese). Kompleksiteten til eukaryote celler tillot dem å organisere en flercellet organisme, i utviklingsprosessen for å gi alt det grunnleggende mangfoldet av liv på jorden.
Tabell over forskjeller mellom prokaryoter og eukaryoter
| skilt | prokaryoter | eukaryoter |
|---|---|---|
| cellekjernen | Ikke | Det er |
| Membranorganeller | Nei. Deres funksjoner utføres av invaginasjoner av cellemembranen, hvor pigmenter og enzymer er plassert. | Mitokondrier, plastider, lysosomer, ER, Golgi-kompleks |
| cellemembraner | Mer komplisert, det er forskjellige kapsler. Celleveggen består av murein. | Hovedkomponenten i celleveggen er cellulose (i planter) eller kitin (i sopp). Dyreceller har ikke cellevegg. |
| genetisk materiale | Betydelig mindre. Det er representert av en nukleoid og plasmider, som har en ringform og er lokalisert i cytoplasmaet. | Mengden arvelig informasjon er betydelig. Kromosomer (som består av DNA og proteiner). preget av diploidi. |
| Inndeling | Binær celledeling. | Det er mitose og meiose. |
| flercellethet | Ikke typisk for prokaryoter. | De er representert av både encellede og flercellede former. |
| Ribosomer | mindre | Større |
| Metabolisme | Mer mangfoldig (heterotrofer, fotosyntetisk og kjemosyntetisk forskjellige måter autotrofer; anaerob og aerob respirasjon). | Autotrofi kun i planter gjennom fotosyntese. Nesten alle eukaryoter er aerobe. |
| Opprinnelse | Fra livløs natur i ferd med kjemisk og prebiologisk evolusjon. | Fra prokaryoter i løpet av deres biologiske evolusjon. |
Den prokaryote cellen er mye enklere enn cellene til dyr og planter. Utvendig er den dekket med en cellevegg som utfører beskyttende, formende og transporterende funksjoner. Stivheten til celleveggen er gitt av murein. Noen ganger er bakteriecellen dekket på toppen med en kapsel eller slimlag.
Protoplasmaet til bakterier, som det til eukaryoter, er omgitt av plasmamembran. Mesosomer involvert i respirasjonsprosessen, bakterioklorofyll og andre pigmenter finnes i sekklignende, rørformede eller lamellære invaginasjoner av membranen. Det genetiske materialet til prokaryoter danner ikke en kjerne, men befinner seg direkte i cytoplasmaet. Bakteriell DNA er et enkelt sirkulært molekyl, som hver består av tusenvis og millioner av basepar. Genomet til en bakteriecelle er mye enklere enn cellene til mer avanserte skapninger: i gjennomsnitt inneholder bakteriell DNA flere tusen gener.
Finnes ikke i prokaryote celler endoplasmatisk retikulum, a ribosomer flyter fritt i cytoplasmaet. Ikke hos prokaryoter mitokondrier; en del av deres funksjon utføres av cellemembranen.
prokaryoter
Bakterier er de minste av organismene med en cellulær struktur; deres størrelse varierer fra 0,1 til 10 µm. Et typisk utskriftssted kan romme hundretusenvis av mellomstore bakterier. Bakterier kan bare sees gjennom et mikroskop, og det er derfor de kalles mikroorganismer eller mikrober; mikroorganismer studeres mikrobiologi . Den delen av mikrobiologien som omhandler bakterier kalles bakteriologi . Begynnelsen på denne vitenskapen lå Anthony van Leeuwenhoek på 1600-tallet.
bakterie er de eldste kjente organismene. Spor av vital aktivitet av bakterier og blågrønnalger (stromatolitter) tilhører arkeene og dateres tilbake til 3,5 milliarder år.På grunn av muligheten for genutveksling mellom representanter forskjellige typer og til og med fødsel, er det ganske vanskelig å systematisere prokaryoter. Tilfredsstillende taksonomi av prokaryoter er ennå ikke bygget; alle eksisterende systemer er kunstige og klassifiserer bakterier i henhold til en gruppe karakterer, uten å ta hensyn til deres fylogenetiske forhold. Tidligere bakterier sammen med sopp og alger inkludert i underriket av lavere planter. Bakterier er for tiden klassifisert som et eget rike av prokaryoter. Det vanligste klassifiseringssystemet er Bergi system basert på strukturen til celleveggen.
På slutten av 1900-tallet oppdaget forskerne at cellene til en relativt lite studert gruppe bakterier - arkebakterier - inneholde rRNA, som skiller seg i struktur fra både prokaryot rRNA og eukaryotisk rRNA. Strukturen til det genetiske apparatet til arkebakterier (tilstedeværelsen introner og repeterende sekvenser behandling, formen ribosom) bringer dem nærmere eukaryoter; på den annen side har arkebakterier også typiske tegn på prokaryoter (fravær av en kjerne i cellen, tilstedeværelse av flageller, plasmider og gassvakuoler, rRNA-størrelse, nitrogenfiksering). Til slutt skiller arkebakterier seg fra alle andre organismer i strukturen til celleveggen, typen fotosyntese og noen andre funksjoner. Arkebakterier er i stand til å eksistere under ekstreme forhold (for eksempel i varme kilder ved temperaturer over 100 ° C, i havdypet ved et trykk på 260 atm, i mettede saltløsninger (30% NaCl)). Noen arkebakterier produserer metan, mens andre bruker svovelforbindelser til energi.
Tilsynelatende er arkebakterier en veldig gammel gruppe organismer; "ekstreme" muligheter vitner om forholdene som er karakteristiske for jordoverflaten i arkeisk tid. Det antas at arkebakterier er nærmest de hypotetiske "cellene" som senere ga opphav til alt mangfoldet av liv på jorden.
Nylig har det blitt klart at det er tre hovedtyper rRNA, presentert henholdsvis den første - i eukaryote celler, den andre - i cellene til ekte bakterier, så vel som i mitokondrier og kloroplaster eukaryoter, den tredje - i arkebakterier. Studier av molekylær genetikk har tvunget frem et nytt blikk på teorien om opprinnelsen til eukaryoter. Det antas for tiden at tre forskjellige grener av prokaryoter utviklet seg samtidig på den gamle jorden - arkebakterier, eubakterier og urkaryoter , preget av en annen struktur og ulike måter å skaffe energi på. Urkaryoter, som faktisk var den nukleære-cytoplasmatiske komponenten i eukaryoter, ble deretter inkludert som symbionter representanter for ulike grupper av eubakterier, som ble til mitokondrier og kloroplaster av fremtidige eukaryote celler.
Dermed er klasserangeringen som ble tildelt tidligere for arkebakterier klart utilstrekkelig. For tiden har mange forskere en tendens til å dele prokaryoter i to riker: arkebakterier og ekte bakterier (eubakterier ) eller til og med skille arkebakterier i et eget kongerike Archaea.
Klassifiseringen av ekte bakterier er gitt i ordningen.
PÅ bakteriecelle det er ingen kjerne, kromosomene er fritt plassert i cytoplasmaet. I tillegg er det ingen membranorganeller i bakteriecellen: mitokondrier, EPS, Golgi-apparatet osv. Utvendig er cellemembranen dekket med en cellevegg.
De fleste bakterier beveger seg passivt, ved hjelp av vann eller luftstrømmer. Bare noen få av dem har bevegelsesorganeller - flagella . Prokaryote flageller er svært enkle i strukturen og består av flagellinproteinet, som danner en hul sylinder på 10–20 nm i diameter. De skrus inn i mediet og beveger cellen fremover. Tilsynelatende er dette den eneste strukturen kjent i naturen som bruker hjulprinsippet.
Bakterier er delt inn i flere grupper i henhold til deres form:
kokker (har en avrundet form);
basiller (ha en stavformet form);
spirilla (ha form som en spiral);
vibrios (har form av komma).
I henhold til åndedrettsmåten er bakterier delt inn i aerober (de fleste bakterier) og anaerobe (årsakene til stivkrampe, botulisme, gass koldbrann). Førstnevnte trenger oksygen for å puste, for sistnevnte oksygen er ubrukelig eller til og med giftig.
Bakterier formerer seg ved å dele seg omtrent hvert 20. minutt (under gunstige forhold). DNA replikerer, hver dattercelle mottar sin egen kopi av foreldre-DNA. Det er også mulig å overføre DNA mellom ikke-delte celler (gjennom fangst av "nakent" DNA, ved hjelp av bakteriofager eller av konjugasjoner , når bakterier er sammenkoblet av kopulatoriske fimbriae), forekommer imidlertid ikke en økning i antall individer. Reproduksjon forhindres av solens stråler og produktene av deres egen vitale aktivitet.Oppførselen til bakterier er ikke spesielt kompleks. Kjemiske reseptorer registrerer endringer i surheten i miljøet og konsentrasjonen av ulike stoffer: sukkerarter, aminosyrer, oksygen. Mange bakterier reagerer på endringer i temperatur eller lys, og noen bakterier kan føle jordas magnetfelt.
Under ugunstige forhold er bakterien dekket med et tett skall, cytoplasmaet er dehydrert, og vital aktivitet stopper nesten. I denne tilstanden kan bakteriesporer holde seg i timevis i et dypt vakuum, tåle temperaturer fra -240 °C til +100 °C.
1. En prokaryot celle er preget av tilstedeværelsen
A) ribosom
B) mitokondrier
B) en dekorert kjerne
D) plasmamembran
D) endoplasmatisk retikulum
E) ett sirkulært DNA
Svar
2. Prokaryote celler er forskjellige fra eukaryote celler
A) tilstedeværelsen av ribosomer
B) mangel på mitokondrier
B) fraværet av en formalisert kjerne
D) tilstedeværelsen av en plasmamembran
D) mangel på bevegelsesorganeller
E) tilstedeværelsen av ett ringkromosom
Svar
3. Etabler samsvar mellom strukturen til celler og deres type: 1-prokaryot, 2-eukaryot
A) ikke har en formalisert kjerne
B) har en kjernemembran
B) diploid eller haploid
D) alltid haploid
D) har ikke mitokondrier, Golgi-komplekset
E) inneholder mitokondrier, Golgi-komplekset
Svar
A1 B2 C2 D1 E1 E2
4. Hvorfor klassifiseres bakterier som prokaryoter?
A) inneholder en kjerne i cellen, isolert fra cytoplasma
B) består av mange differensierte celler
B) har ett ringkromosom
D) har ikke et cellesenter, Golgi-kompleks og mitokondrier
D) ikke har en kjerne isolert fra cytoplasma
E) har cytoplasma og plasmamembran
Svar
5. En bakteriecelle er klassifisert som en prokaryot celle, siden den
A) har ikke en kjerne dekket med et skall
B) har et cytoplasma
B) har ett DNA-molekyl innebygd i cytoplasmaet
D) har en ytre plasmamembran
D) har ikke mitokondrier
E) har ribosomer der proteinsyntese finner sted
Svar
6. Celler av eukaryote organismer, i motsetning til prokaryote, har
A) cytoplasma
B) kjerne dekket med et skall
B) DNA-molekyler
D) mitokondrier
D) hardt skall
E) endoplasmatisk retikulum
Svar
7. Etablere samsvar mellom egenskapene til cellen og dens type: 1-prokaryotisk, 2-eukaryot
A) Membranorganeller er fraværende
B) Det er en cellevegg av murein
C) Arvestoff er representert ved en nukleoid
D) Inneholder kun små ribosomer
E) Arvemateriale er representert ved lineært DNA
E) Cellulær respirasjon forekommer i mitokondrier
Svar
A1 B1 C1 D1 E2 E2
8. Prokaryote celler er forskjellige fra eukaryote celler
A) tilstedeværelsen av en nukleoid i cytoplasmaet
B) tilstedeværelsen av ribosomer i cytoplasmaet
C) ATP-syntese i mitokondrier
D) tilstedeværelsen av det endoplasmatiske retikulum
D) fraværet av en morfologisk distinkt kjerne
E) tilstedeværelsen av invaginasjoner av plasmamembranen, som utfører funksjonen til membranorganeller
I henhold til cellens struktur er levende organismer delt inn i prokaryoter og eukaryot. Celler av begge er omgitt plasmamembran, utenfor som det i mange tilfeller er celleveggen. Inne i cellen er en halvvæske cytoplasma. Imidlertid er prokaryote celler mye enklere enn eukaryote celler.
Grunnleggende genetisk materiale prokaryoter (fra gresk. Om- før og karyon- kjerne) ligger i cytoplasmaet i form av et sirkulært DNA-molekyl. Dette molekylet ( nukleoid) er ikke omgitt av en kjernemembran som er karakteristisk for eukaryoter, og er festet til plasmamembranen (fig. 1). Dermed har ikke prokaryoter en velformet kjerne. I tillegg til nukleoiden finnes ofte et lite sirkulært DNA-molekyl i en prokaryot celle, kalt plasmid. Plasmider kan bevege seg fra en celle til en annen og integreres i hoved-DNA-molekylet.
Noen prokaryoter har utvekster av plasmamembranen: mesosomer, lamellære thylakoider, kromatoforer. De inneholder enzymer involvert i fotosyntese og respirasjon. I tillegg er mesosomer assosiert med DNA-syntese og proteinsekresjon.
Prokaryote celler er små, deres diameter er 0,3-5 mikron. På utsiden av plasmamembranen til alle prokaryoter (med unntak av mycoplasmas) er celleveggen. Den består av komplekser av proteiner og oligosakkarider, stablet i lag, beskytter cellen og opprettholder formen. Den er atskilt fra plasmamembranen med et lite mellomrom.
Bare ikke-membranorganeller finnes i cytoplasmaet til prokaryoter. ribosomer. Strukturen til ribosomer av prokaryoter og eukaryoter er lik, men ribosomer av prokaryoter er mindre og er ikke festet til membranen, men er lokalisert direkte i cytoplasma.

Mange prokaryoter er bevegelige og kan svømme eller gli ved hjelp av flageller.
Prokaryoter formerer seg vanligvis ved fisjon i to ( binær). Delingen innledes av et veldig kort stadium med dobling eller replikering av kromosomer. Så prokaryoter er haploide organismer.
Prokaryoter inkluderer bakterier og blågrønne alger, eller cyanobakterier. Prokaryoter dukket opp på jorden for rundt 3,5 milliarder år siden og var sannsynligvis den første cellulære livsformen, og ga opphav til moderne prokaryoter og eukaryoter.
eukaryoter (fra gresk. eu- sant, karyon- kjerne), i motsetning til prokaryoter, har en dannet kjerne omgitt av kjernefysisk konvolutt- dobbeltlags membran. DNA-molekylene som finnes i kjernen er ikke lukkede (lineære molekyler). I tillegg til kjernen, finnes en del av den genetiske informasjonen i DNAet til mitokondrier og kloroplaster. Eukaryoter dukket opp på jorden for rundt 1,5 milliarder år siden.
I motsetning til prokaryoter, representert av enkeltorganismer og koloniale former, kan eukaryoter være encellede (for eksempel amøbe), koloniale (volvox) og flercellede organismer. De er delt inn i tre store riker: dyr, planter og sopp.
Diameteren til eukaryote celler er 5–80 µm. Som prokaryote celler er eukaryote celler omgitt av plasmamembran sammensatt av proteiner og lipider. Denne membranen fungerer som en selektiv barriere, permeabel for noen forbindelser og ugjennomtrengelig for andre. Utenfor plasmamembranen er en sterk celleveggen, som i planter hovedsakelig består av cellulosefibre, og i sopp - av kitin. Hovedfunksjonen til celleveggen er å sikre den konstante formen til cellene. Siden plasmamembranen er permeabel for vann, og plante- og soppceller vanligvis kommer i kontakt med løsninger med lavere ionestyrke enn ionestyrken til løsningen inne i cellen, vil vann komme inn i cellene. På grunn av dette vil cellevolumet øke, plasmamembranen vil begynne å strekke seg og kan gå i stykker. Celleveggen forhindrer celleutvidelse og ødeleggelse.
Hos dyr er celleveggen fraværende, men det ytre laget av plasmamembranen er beriket med karbohydratkomponenter. Dette ytre laget av plasmamembranen til dyreceller kalles glykokalyx. Celler fra flercellede dyr trenger ikke en sterk cellevegg, siden det er andre mekanismer som sikrer regulering av cellevolum. Siden cellene til flercellede dyr og encellede organismer lever i havet er i et miljø der den totale konsentrasjonen av ioner er nær den intracellulære konsentrasjonen av ioner, cellene sveller ikke og sprekker ikke. Encellede dyr som lever i ferskvann (amoeba, ciliate sko) har kontraktile vakuoler som stadig får ut vannet som kommer inn i cellen.
Strukturelle komponenter i en eukaryot celle
Inne i cellen under plasmamembranen er cytoplasma. Hovedstoffet i cytoplasmaet (hyaloplasma) er en konsentrert løsning av uorganiske og organiske forbindelser, hvis hovedkomponenter er proteiner. den kolloidsystem, som kan endres fra flytende til geltilstand og omvendt. En betydelig del av de cytoplasmatiske proteinene er enzymer som utfører ulike kjemiske reaksjoner. ligger i hyaloplasmaet organeller, utføre ulike funksjoner i cellen. Organeller kan være membran (kjerne, Golgi-apparat, endoplasmatisk retikulum, lysosomer, mitokondrier, kloroplaster) og ikke-membran (cellesenter, ribosomer, cytoskjelett).
Membranorganellerhovedkomponenten i membranorganeller er membran. Biologiske membraner bygges iht generelt prinsipp, men kjemisk oppbygning membraner av forskjellige organeller er forskjellige. Alle cellemembraner er tynne filmer (7–10 nm tykke), som er basert på et dobbeltlag av lipider (bilag) arrangert slik at de ladede hydrofile delene av molekylene kommer i kontakt med mediet, og de hydrofobe restene. fettsyrer av hvert monolag er rettet inn i membranen og kommer i kontakt med hverandre (fig. 3). Proteinmolekyler (integrerte membranproteiner) bygges inn i lipid-dobbeltlaget på en slik måte at de hydrofobe delene av proteinmolekylet kommer i kontakt med fettsyrerestene i lipidmolekylene, og de hydrofile delene utsettes for miljøet. I tillegg er en del av de løselige (ikke-membranproteinene) koblet til membranen hovedsakelig på grunn av ioniske interaksjoner (perifere membranproteiner). Karbohydratfragmenter er også festet til mange proteiner og lipider i sammensetningen av membraner. På denne måten, biologiske membraner er lipidfilmer der integrerte proteiner er innebygd.
En av hovedfunksjonene til membraner er å skape en grense mellom cellen og miljøet og cellens ulike rom. Lipid-dobbeltlaget er gjennomtrengelig hovedsakelig for fettløselige forbindelser og gasser, hydrofile stoffer transporteres gjennom membraner ved hjelp av spesielle mekanismer: lav molekylvekt - ved hjelp av en rekke bærere (kanaler, pumper, etc.), og høy molekylvekt - ved hjelp av prosesser exo- og endocytose(Fig. 4).

Ris. 4. Skjema for overføring av stoffer gjennom membranen
På endocytose visse stoffer sorberes på membranoverflaten (på grunn av interaksjon med membranproteiner). På dette stedet dannes en invaginasjon av membranen inn i cytoplasmaet. Deretter separeres en vesikkel fra membranen, inne i hvilken den overførte forbindelsen er inneholdt. På denne måten, endocytose er transport av høymolekylære forbindelser inn i cellen eksternt miljø omgitt av en del av membranen. omvendt prosess, det er eksocytose er transport av stoffer fra cellen til utsiden. Det skjer ved fusjon med plasmamembranen til en vesikkel fylt med transportert makromolekylære forbindelser. Vesikkelmembranen smelter sammen med plasmamembranen, og innholdet helles ut.
Kanaler, pumper og andre transportører er integrerte membranproteinmolekyler som vanligvis danner en pore i membranen.
I tillegg til funksjonene med å dele rom og gi selektiv permeabilitet, er membraner i stand til å oppfatte signaler. Denne funksjonen utføres av reseptorproteiner som binder signalmolekyler. Individuelle membranproteiner er enzymer som utfører visse kjemiske reaksjoner.
Cellekjernen - et stort organell av cellen, omgitt av en kjernemembran og vanligvis har en sfærisk form. Det er bare én kjerne i cellen, og selv om det er flerkjernede celler (celler i skjelettmuskulatur, noen sopp) eller de uten kjerne (erytrocytter og blodplater fra pattedyr), oppstår disse cellene fra mononukleære stamceller.
Hovedfunksjonen til kjernen er lagring, overføring og salg av genetisk informasjon. Her dupliseres DNA-molekyler, som et resultat av at datterceller ved deling mottar det samme genetiske materialet. I kjernen, ved å bruke individuelle seksjoner av DNA-molekyler (gener) som en matrise, syntetiseres RNA-molekyler: informasjons- (mRNA), transport- (tRNA) og ribosomale (rRNA) molekyler som er nødvendige for proteinsyntese. I kjernen er ribosomunderenheter satt sammen av rRNA-molekyler og proteiner, som syntetiseres i cytoplasmaet og overføres til kjernen.
Kjernen består av kjernemembranen, kromatin (kromosomer), nukleolus og nukleoplasma (karyoplasma).

Ris. 5. Struktur av kromatin: 1 - nukleosom, 2 - DNA
Under et mikroskop er soner med tett stoff synlige inne i kjernen - kromatin. I ikke-delte celler fyller den jevnt ut volumet av kjernen eller kondenserer på separate steder i form av tettere områder og flekker godt med grunnleggende fargestoffer. Kromatin er et kompleks av DNA og proteiner (fig. 5), for det meste positivt ladet histoner.
Antall DNA-molekyler i kjernen er lik antall kromosomer. Antallet og formen på kromosomer er et unikt kjennetegn ved arten. Hvert kromosom inneholder ett DNA-molekyl som består av to sammenkoblede tråder og har form av en dobbel helix 2 nm tykk. Lengden overstiger cellens diameter betydelig: den kan nå flere centimeter. DNA-molekylet er negativt ladet, så det kan foldes (kondensere) først etter binding til positivt ladede histonproteiner (fig. 6).
Først vrir den doble DNA-strengen seg rundt separate blokker av histoner, som hver inkluderer 8 proteinmolekyler, og danner en struktur i form av "perler på en streng" med en tykkelse på omtrent 10 nm. Perlene kalles nukleosomer. Som et resultat av dannelsen av nukleosomer reduseres lengden på DNA-molekylet med omtrent 7 ganger. Deretter kveiles tråden med nukleosomer, og danner en struktur i form av et tau med en tykkelse på omtrent 30 nm. Deretter festes et slikt tau, bøyd i form av løkker, til proteinene som danner grunnlaget for kromosomet. Som et resultat dannes en struktur med en tykkelse på omtrent 300 nm. Ytterligere kondensering av denne strukturen fører til dannelsen av et kromosom.
Mellom delingene utfolder kromosomet seg delvis. Som et resultat blir individuelle deler av DNA-molekylet som skal uttrykkes i en gitt celle frigjort fra proteiner og strukket, noe som gjør det mulig å lese informasjon fra dem ved å syntetisere RNA-molekyler.
Nukleolus er en type mal DNA som er ansvarlig for syntesen av rRNA og er satt sammen i separate områder av kjernen. Nucleolus er den tetteste strukturen i kjernen; det er ikke en separat organell, men er en av lokiene til kromosomet. Den produserer rRNA, som deretter danner et kompleks med proteiner, og danner ribosomale underenheter som går inn i cytoplasmaet.
Ikke-histonproteinene i kjernen danner et strukturelt nettverk i kjernen. Det er representert av et lag med fibriller som ligger under kjernefysiske konvolutten. Et intranukleært nettverk av fibriller er festet til det, som kromatinfibriller er festet til.
Den kjernefysiske konvolutten består av to membraner: ytre og indre, atskilt av et intermembranrom. Den ytre membranen er i kontakt med cytoplasma, den kan inneholde polyribosomer, og den kan passere inn i membranene i det endoplasmatiske retikulum. Den indre membranen er assosiert med kromatin. Dermed sikrer kjernefysisk konvolutt fiksering av kromosomalt materiale i det tredimensjonale rommet til kjernen.
Skallet til kjernen har runde hull - kjernefysiske porer(Fig. 7). I området av poren lukkes de ytre og indre membranene og danner hull fylt med fibriller og granulat. Inne i porene er et komplekst system av proteiner som gir selektiv binding og overføring av makromolekyler. Antall kjernefysiske porer avhenger av intensiteten av cellemetabolismen.
Endoplasmatisk retikulum, eller endoplasmatisk retikulum (EPR), er et bisarrt nettverk av kanaler, vakuoler, flatede sekker, sammenkoblet og atskilt fra hyaloplasmaet med en membran (fig. 8).
Skille ujevn og glatt EPJ . På membranene til den grove ER er ribosomer(Fig. 9), som syntetiserer proteiner som skilles ut fra cellen eller inkorporeres i plasmamembranen. Det nylig syntetiserte proteinet forlater ribosomet og passerer gjennom en spesiell kanal inn i hulrommet endoplasmatisk retikulum, hvor den gjennomgår post-translasjonell modifikasjon, for eksempel binding til karbohydrater, proteolytisk spaltning av en del av polypeptidkjeden, dannelse av S–S-bindinger mellom cysteinrester i kjeden. Videre transporteres disse proteinene til Golgi-komplekset, hvor de enten er en del av lysosomer eller sekretoriske granuler. I begge tilfeller er disse proteinene inne i membranvesikkelen (vesikkel).
Ris. Fig. 9. Skjema for proteinsyntese i en grov ER: 1 – liten og
2 - stor underenhet av ribosomet; 3 - rRNA-molekyl;
4 - grov EPJ; 5 - nylig syntetisert protein
Den glatte ER mangler ribosomer. Dens hovedfunksjon er lipidsyntese og karbohydratmetabolisme. Det er godt utviklet, for eksempel i cellene i binyrebarken, som inneholder enzymer som sørger for syntese av steroidhormoner. Den glatte ER i leverceller inneholder enzymer som oksiderer (avgifter) hydrofobe forbindelser som er fremmede for kroppen, for eksempel medikamenter.

Ris. 10. Golgi-apparat: 1 - vesikler; 2 - tanker
Golgi kompleks (Fig. 10) består av 5–10 flate membranbundne hulrom anordnet parallelt. Endedelene av disse skiveformede strukturene har forlengelser. Det kan være flere slike formasjoner i en celle. I sonen av Golgi komplekset er et stort nummer av membranvesikler. Noen av dem er snøret fra endedelene av hovedstrukturen i form av sekretoriske granuler og lysosomer. Noen av de små vesiklene (vesiklene) som bærer proteiner syntetisert i den grove ER flytter til Golgi-komplekset og smelter sammen med det. Dermed er Golgi-komplekset involvert i akkumulering og videre modifisering av produkter syntetisert i grov EPR og deres sortering.

Ris. 11. Dannelse og funksjoner av lysosomer: 1 - fagosom; 2 - pinocytisk vesikkel; 3 - primær lysosom; 4 - Golgi-apparater; 5 - sekundær lysosom
Lysosomer - dette er vakuoler (fig. 11), begrenset av én membran, som knopper fra Golgi-komplekset. Det er nok lysosomer inni surt miljø(pH 4,9–5,2). Det er hydrolytiske enzymer som bryter ned ulike polymerer ved sur pH (proteaser, nukleaser, glukosidaser, fosfataser, lipaser). Disse primære lysosomer smelter sammen med endocytiske vakuoler som inneholder komponenter som skal spaltes. Stoffer som har kommet inn i det sekundære lysosomet brytes ned til monomerer og overføres gjennom lysosommembranen til hyaloplasmaet. Dermed er lysosomer involvert i prosessene med intracellulær fordøyelse.
Mitokondrier omgitt av to membraner: den ytre, som skiller mitokondriene fra hyaloplasma, og den indre, som avgrenser dets indre innhold. Mellom dem er det et intermembranrom 10–20 nm bredt. Den indre membranen danner mange utvekster ( cristae). Denne membranen inneholder enzymer som sikrer oksidasjon av aminosyrer, sukkerarter, glyserol og fettsyrer dannet utenfor mitokondriene (Krebs-syklusen) og utfører elektronoverføring i respirasjonskjeden (skjemaet). På grunn av overføring av elektroner langs respirasjonskjeden fra et høyt til et lavere energinivå, lagres en del av den frigjorte frie energien i form av ATP, cellens universelle energivaluta. Dermed er hovedfunksjonen til mitokondrier oksidasjon av forskjellige substrater og syntese av ATP-molekyler.
Skjema for overføring av to elektroner langs respirasjonskjeden
Inne i mitokondriene er et sirkulært DNA-molekyl som koder for noen av mitokondrieproteinene. I det indre rommet av mitokondrier (matrise) er det ribosomer som ligner på prokaryoter, som gir syntesen av disse proteinene.
Det faktum at mitokondrier har sitt eget sirkulære DNA og prokaryote ribosomer har ført til hypotesen om at mitokondriet er en etterkommer av en eldgammel prokaryot celle som en gang kom inn i den eukaryote cellen og tok på seg separate funksjoner i evolusjonsprosessen.
Ris. 12. Kloroplaster (A) og thylakoidmembraner (B)
plastider – organeller Plante-celle som inneholder pigmenter. PÅ kloroplaster Inneholder klorofyll og karotenoider kromoplaster- karotenoider, leukoplaster det er ingen pigmenter. Plastider er omgitt av en dobbel membran. Inne i dem er et system av membraner i form av flate vesikler kalt thylakoider(Fig. 12). Thylakoider er stablet i stabler som ligner stabler av plater. Pigmenter er innebygd i thylakoidmembraner. Deres hovedfunksjon er å absorbere lys, hvis energi, ved hjelp av enzymer innebygd i tylakoidmembranen, omdannes til en gradient av H + -ioner på tylakoidmembranen. Som mitokondrier har plastider sitt eget sirkulære DNA og ribosomer av prokaryot-typen. Tilsynelatende er plastider også en prokaryot organisme som lever i symbiose med eukaryote celler.
Ribosomer er ikke-membrancelleorganeller som finnes i både pro- og eukaryote celler. Eukaryote ribosomer er større enn prokaryote, størrelsen deres er 25x20x20 nm. Ribosomet består av store og små underenheter ved siden av hverandre. En mRNA-streng er plassert mellom underenhetene i et fungerende ribosom.
Hver underenhet av ribosomet er bygget fra rRNA, tettpakket og assosiert med proteiner. Ribosomer kan lokaliseres fritt i cytoplasmaet eller assosieres med ER-membraner. Frie ribosomer kan være enkeltstående, men kan danne polysomer når flere ribosomer er sekvensielt plassert på en mRNA-streng. Hovedfunksjonen til ribosomer er proteinsyntese.
cytoskjelett - dette er muskel- og skjelettsystemet celler, inkludert proteinfilamentøse (fibrillære) formasjoner, som er skjelettet til cellen og utfører motorisk funksjon. Strukturene i cytoskjelettet er dynamiske, de oppstår og går i oppløsning. Cytoskjelettet er representert av tre typer formasjoner: mellomliggende filamenter(filamenter med en diameter på 10 nm), mikrofilament s (filamenter med en diameter på 5–7 nm) og mikrotubuli. Mellomfilamenter er ikke-forgrenende proteinstrukturer i form av filamenter, ofte arrangert i bunter. Dem proteinsammensetning forskjellige i forskjellige vev: i epitelet består de av keratin, i fibroblaster - fra vimentin, i muskelceller- fra desmin. Mellomliggende filamenter vil utføre en støtterammefunksjon.
Mikrofilamenter - dette er fibrillære strukturer som ligger rett under plasmamembranen i form av bunter eller lag. De er tydelig synlige i amøbens prolegs, i bevegelsesprosessene til fibroblaster og i mikrovilli i tarmepitelet (fig. 13). Mikrofilamenter er bygget opp fra de kontraktile proteinene aktin og myosin og er et intracellulært kontraktilt apparat.
mikrotubuli er en del av både midlertidige og permanente strukturer i cellen. De temporale inkluderer delingsspindelen, elementer i cytoskjelettet til celler mellom delingene, og de permanente inkluderer flimmerhår, flageller og centrioler i cellesenteret. Mikrotubuli er rette hule sylindre med en diameter på omtrent 24 nm, veggene deres er dannet av avrundede tubulinproteinmolekyler. Under et elektronmikroskop kan man se at mikrotubuli-tverrsnittet er dannet av 13 underenheter koblet i en ring. Mikrotubuli er tilstede i hyaloplasmaet til alle eukaryote celler. En av funksjonene til mikrotubuli er å lage et stillas inne i cellene. I tillegg beveger små vesikler seg langs mikrotubuli, som på skinner.
Cellesenter består av to sentrioler plassert i rette vinkler på hverandre og tilhørende mikrotubuli. Disse organellene i delende celler deltar i dannelsen av delingsspindelen. Grunnlaget for sentriolen er 9 tripletter av mikrotubuli plassert rundt omkretsen, og danner en hul sylinder, 0,2 µm bred og 0,3–0,5 µm lang. Som forberedelse til celledeling separeres sentriolene og dobles. Før mitose er sentrioler involvert i dannelsen av spindelmikrotubuli. Høyere planteceller har ikke sentrioler, men de har et lignende organiseringssenter for mikrotubulus.